第一节 从单细胞到多细胞
在动物界里除了单细胞动物外,其余都是多细胞动物。从单细胞到多细胞是生物从低级向高级发展的一个重要过程,代表了生物进化史上一个极为重要的阶段。一切高等生物虽然都是多细胞的,但发展是不平衡的。动物的发展水平远远高于植物,它们进化发展的速度也远较植物为快。动物的基本特点之一是有对称的体型。两侧对称的体型不仅有利于活动,且促使身体分为前后、左右和背腹。在进化过程中,神经感官和取食器官逐渐向前端集中,形成了头部。对称体型和头部的形成是动物体复杂化的关键。一切高等动物以至于人都是在这一体型基础上发展起来的。
单细胞动物在形态结构上虽然有的也较复杂,但它只是一个细胞本身的分化。它们之中虽然也有群体,但是群体中的每个个体细胞,一般还是独立生活,彼此间的联系并不密切,因此,在发展上它们是处于低级的、原始阶段,属于原生动物。
绝大多数多细胞动物叫做后生动物(Metazoa),这和原生动物的名称是相对而言的。
在原生动物和后生动物之间,长期以来学者们认为还有一类中生动物(Mesozoa)(图3-1),中生动物这个名字就是因为,认为中生动物介于原生动物和后生动物之间。有学者将原生动物、中生动物、后生动物并列为3个动物亚界。现在一般认为中生动物为动物界中的一门。中生动物是一类小型的内寄主动物。结构简单,已知约50种,分为菱形虫纲(Rhombozoa)和直泳虫纲(Orthonecta),前者包括双胚虫(dicyemida)和异胚虫(heterocyemida)两类。菱形虫纲的动物寄生在头足类软体动物的肾内,体长约0.5mm~10mm,虫体由20~40个细胞组成,细胞数目在每个种内是恒定的。这些细胞基本上排列成双层,但又不同于高等动物的胚层。外层是单层具纤毛的体细胞,包围着中央的一个或几个延长的轴细胞。虫体前端的8~9个体细胞排成两圈,用以附着寄主。其余的体细胞多少呈螺旋形排列(图3—1)。体细胞具营养的功能,轴细胞具繁殖功能。同无性生殖和有性生殖(图3—1)。生活史较为复杂,尚不完全了解。直泳虫纲的动物寄生在多种海生无脊椎动物体内(如扁形动物、纽形动物、环节动物、双壳贝类及棘皮动物)。成虫多数雌雄异体(图3—1C),雌性个体较雄性大,外层亦为单层具纤毛的体细胞,呈环形整齐排列,前端体细胞的纤毛指向前方,其余的纤毛向后方,体细胞中央围绕着许多生殖细胞(卵或精子)。少数种类,成虫雌雄同体,其精细胞在卵细胞的前方。没有轴细胞。性成熟后,雄性个体释放精子到海水中,精子进入雌性个体内与卵受精,并在雌体内发育成具纤毛的幼虫(一层纤毛细胞包围几个生殖细胞)(图3—1)。幼虫离开母体又感染新寄主。当幼虫侵入寄主组织,其外层具纤毛的细胞消失,生殖细胞多分裂形成多核的变形体(plasmodium)(图3—2)。变形体由无性的碎裂方法产生很多变形体,然后由它们发育成雌、雄个体。
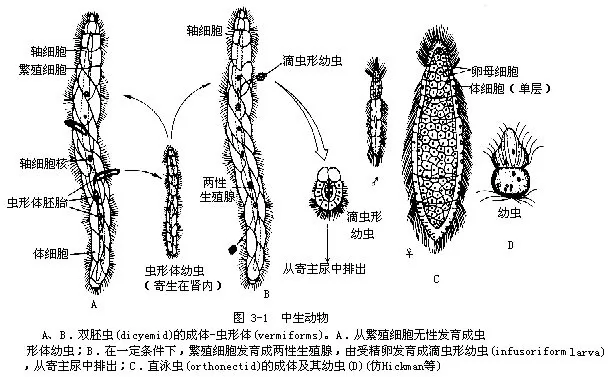
近十余年来对中生动物的系统发育、亚显微结构、生理、生殖、发育、生态以及生化分类等进行了多方面的研究。目前对中生动物的系统发育关系仍存在着争议。有些学者基于中生动物全部为寄生,且生活史较复杂,结构简单是适应寄生生活的退化现象,因此认为它是退化的扁形动物。甚至认为可以作为一纲列入扁形动物门。还有一些学者基于其身体结构有体细胞和生殖细胞的分化,体表具纤毛,且其寄生历史较长,因此认为中生动物是原始的种类,是由最原始的多细胞动物进化来的,或认为是早期后生动物的一个分支。近年来经生化分析表明,中生动物细胞核DNA中鸟嘌呤和胞嘧啶的含量(23%)与原生动物纤毛虫类的含量相近,而低于其它多细胞动物者,包括扁形动物者(35%~50%)。因此认为中生动物和原生动物的纤毛虫类的亲缘关系较近,更可能是真正原始的多细胞动物。至于中生动物和后生动物是否各自独立地来于原生动物的祖先,或中生动物确是原始的或退化的扁虫?还不很清楚。由于中生动物有着长期的寄生历史,是动物界中极为特殊的类群,其分类地位尚难确定。

第二节 多细胞动物起源于单细胞动物的证据
一般公认多细胞动物起源于单细胞动物。其证据是:
(一)古生物学方面 古代动、植物的遗体或遗迹,经过千百万年地壳的变迁或造山运动等,被埋在地层中形成了化石。已经发现在最古老的地层中,化石种类也是最简单的。在太古代的地层中有大量有孔虫壳化石,而在晚近的地层中动物的化石种类也较复杂,并且能看出生物由低等向高等发展的顺序。说明最初出现单细胞动物,后来才发展出多细胞动物。从辩证唯物主义的观点来看,事物的发展是由简单到复杂、由低等到高等,生物的发展也不例外。
(二)形态学方面 从现有动物来看,有单细胞动物、多细胞动物,并形成了由简单到复杂、由低等到高等的序列。在原生动物鞭毛纲中有些群体鞭毛虫,如团藻,其形态与多细胞动物很相似,可推测这类动物是从单细胞动物过渡到多细胞动物的中间类型,即由单细胞动物发展成群体以后,又进一步发展成多细胞动物。
(三)胚胎学方面 在胚胎发育中,多细胞动物是由受精卵开始,经过卵裂、囊胚、原肠胚等一系列过程,逐渐发育成成体。多细胞动物的早期胚胎发育基本上是相似的。根据生物发生律,个体发育简短地重演了系统发展的过程,可以说明多细胞动物起源于单细胞动物,并且说明多细胞动物发展的早期所经历的过程是相似的。恩格斯说:“有机体的胚胎向成熟的有机体的逐步发育同植物和动物在地球历史上相继出现的次序之间有特殊的吻合。正是这种吻合为进化论提供了最可靠的根据。”
第三节 胚胎发育的重要阶段
多细胞动物的胚胎发育比较复杂。不同类的动物,胚胎发育的情况不同,但是早期胚胎发育的几个主要阶段是相同的。
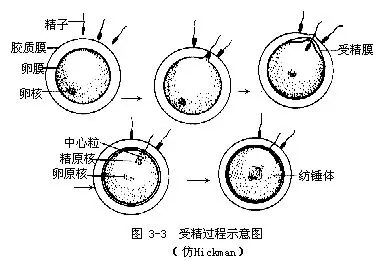
(一)受精(fertilization)与受精卵 由雌、雄个体产生雌雄生殖细胞,雌性生殖细胞称为卵。卵细胞较大,里面一般含有大量卵黄。根据卵黄多少可将卵分为少黄卵、中黄卵和多黄卵。卵黄相对多的一端称为植物极(vegetal pole),另一端称为动物极(animal pole)。雄性生殖细胞称为精子,精子个体小,能活动。精子与卵结合为一个细胞称为受精卵,这个过程就是受精(图3—3)。受精卵是新个体发育的起点,由受精卵发育成新个体。
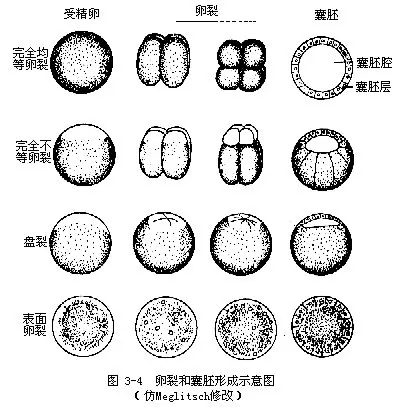
(二)卵裂(cleavage) 受精卵进行卵裂,它与一般细胞分裂的不同点在于每次分裂之后,新的细胞未长大,又继续进行分裂,因此分裂成的细胞越来越小。这些细胞也叫分裂球(blas- tomere)。由于不同类动物卵细胞内卵黄多少及其在卵内分布情况的不同,卵裂的方式也不同:
1.完全卵裂(total cleavage) (图3-4)整个卵细胞都进行分裂,多见于少黄卵。卵黄少、分布均匀,形成的分裂球大小相等的叫等裂,如海胆、文昌鱼。如果卵黄在卵内分布不均匀,形成的分裂球大小不等的叫不等裂,如海绵动物、蛙类。
2.不完全卵裂(partial cleavage) (图 3-4)多见于多黄卵。卵黄多,分裂受阻,受精卵只在不含卵黄的部位进行分裂。分裂区只限于胚盘处的称为盘裂(discal cleavage),如乌贼、鸡卵。分裂区只限于卵表面的称为表面卵裂(peripheral cleavage),如昆虫卵。各种卵裂的结果,其形态虽有差别,但都进入下一发育阶段。
(三)囊胚的形成(blastulation) 卵裂的结果,分裂球形成中空的球状胚,称为囊胚(blastu-la)(图3—4)。囊胚中间的腔称为囊胚腔(blastocoel),囊胚壁的细胞层称为囊胚层(blastoderm)。
(四)原肠胚的形成(gastrulation) 囊胚进一步发育进入原肠胚形成阶段,此时胚胎分化出内、外两胚层和原肠腔。原肠胚形成在各类动物有所不同,其方式有:
1.内陷(invagination)由囊胚植物极细胞向内陷入。最后形成2层细胞,在外面的细胞层称为外胚层(ectoderm),向内陷入的一层为内胚层(endoderm)。内胚层所包围的腔,将形成未来的肠腔,因此称为原肠腔(gastrocoel)。原肠腔与外界相通的孔称为原口或胚孔(blastopore)(图3—5)。
2.内移(ingression)是由囊胚一部分细胞移入内部形成内胚层。开始移入的细胞充填于囊胚腔内,排列不规则,接着逐渐排成一层内胚层。有的移入时就排列成内胚层。这样的原肠胚没有孔,以后在胚的一端开一胚孔(图3—5)。

3.分层(delamination)囊胚的细胞分裂时,细胞沿切线方向分裂,这样向着囊胚腔分裂出的细胞为内胚层,留在表面的一层为外胚层(图3—5)。
4.内转(involution)通过盘裂形成的囊胚,分裂的细胞由下面边缘向内转,伸展成为内胚层(图3—5)。
5.外包(epiboly)动物极细胞分裂快,植物极细胞由于卵黄多分裂极慢,结果动物极细胞逐渐向下包围植物极细胞,形成为外胚层,被包围的植物极细胞为内胚层(图3—5)。
以上原肠胚形成的几种类型常常综合出现,最常见的是内陷与外包同时进行,分层与内移相伴而行。
(五)中胚层及体腔的形成绝大多数多细胞动物除了内、外胚层之外,还进一步发育,在内外胚层之间形成中胚层(mesoderm)。在中胚层之间形成的腔称为真体腔。主要由以下方式形成:
1.端细胞法在胚孔的两侧,内、外胚层交界处各有一个细胞分裂成很多细胞,形成索状,伸入内、外胚层之间,是为中胚层细胞。在中胚层之间形成的空腔即为体腔(真体腔)。由于这种体腔是在中胚层细胞之间裂开形成的,因此又称为裂体腔(schizocoel),这样形成体腔的方式又称为裂体腔法(schizocoelous method或schizocoelic formation)原口动物都是以端细胞法形成中胚层和体腔(图3—6)。

2.体腔囊法在原肠背部两侧,内胚层向外突出成对的囊状突起称体腔囊。体腔囊和内胚层脱离后,在内外胚层之间逐步扩展成为中胚层,由中胚层包围的空腔称为体腔(图3—6)。因为体腔囊来源于原肠背部两侧,所以又称为肠体腔(enterocoel)。这样形成体腔的方式称为肠体腔法(enterocoelous method或enterocoelicformation)。后口动物的棘皮动物、毛颚动物、半索动物及脊索动物均以这种方式形成中胚层和体腔。高等脊索动物是由裂体腔法形成体腔,但具体的形成过程比较复杂,各个类群之间的发育细节也有差异。
(六)胚层的分化胚胎时期的细胞,开始出现时,相对地说是较简单、均质和具有可塑性。进一步发育,由于遗传性、环境、营养、激素以及细胞群之间相互诱导等因素的影响,而转变为较复杂、异质性和稳定性的细胞。这种变化现象称为分化(differentiation)。动物体的组织、器官都是从内、中、外三胚层发育分化而来的。如内胚层分化为消化管的大部分上皮、肝、胰、呼吸器官,排泄与生殖器官的小部分。中胚层分化为肌肉、结缔组织(包括骨骼、血液等)、生殖与排泄器官的大部分。外胚层分化为皮肤上皮(包括上皮各种衍生物如皮肤腺、毛、角、爪等)、神经组织、感觉器官、消化管的两端。
第四节 生物发生律
生物发生律(biogenetic law)也叫重演律(recapitulation law),是德国人赫克尔(E.Haeckel,1834—1919)用生物进化论的观点总结了当时胚胎学方面的工作提出来的。当时在胚胎发育方面已揭示了一些规律,如在动物胚胎发育过程中,各纲脊椎动物的胚胎都是由受精卵开始发育的,在发育初期极为相似,以后才逐渐变得越来越不相同。达尔文用进化论的观点曾作过一些论证,认为胚胎发育的相似性,说明它们彼此有亲缘关系,起源于共同的祖先,个体发育的渐进性是系统发展中渐进性的表现。达尔文还指出了胚胎结构重演其过去祖先的结构,“它重演了它们祖先发育中的一个形象”。
赫克尔明确地论述了生物发生律。1866年他在《普通形态学》一书中是这样说的:“生物发展史可分为2个相互密切联系的部分,即个体发育(ontogeny)和系统发展(或系统发育phyloge-ny),也就是个体的发育历史和由同一起源所产生的生物群的发展历史。个体发育史是系统发展史的简单而迅速的重演。”如青蛙的个体发育,由受精卵开始,经过囊胚、原肠胚、三胚层的胚、无腿蝌蚪、有腿蝌蚪,到成体青蛙。这反映了它在系统发展过程中经历了像单细胞动物、单细胞的球状群体、腔肠动物、原始三胚层动物、鱼类动物,发展到有尾两栖到无尾两栖动物的基本过程。说明了蛙个体发育重演了其祖先的进化过程,也就是个体发育简短重演了它的系统发展,即其种族发展史。
生物发生律对了解各动物类群的亲缘关系及其发展线索极为重要。因而对许多动物的亲缘关系和分类位置不能确定时,常由胚胎发育得到解决。生物发生律是一条客观规律,它不仅适用于动物界,而且适用于整个生物界,包括人在内。当然不能把“重演”理解为机械的重复,而且在个体发育中也会有新的变异出现,个体发育又不断的补充系统发展。这二者的关系是辩证统一的,二者相互联系、相互制约,系统发展通过遗传决定个体发育,个体发育不仅简短重演系统发展,而且又能补充和丰富系统发展。
第五节 关于多细胞动物起源的学说
多细胞动物起源于单细胞动物,至于是哪一类单细胞动物发展成多细胞动物,以及多细胞动物起源的方式如何,有不同学说。
(一)群体学说(colonial theory)认为后生动物来源于群体鞭毛虫,这是后生动物起源的经典学说。有一些日益增多的证据,因而是当代动物学中最广泛接受的学说。这一学说是由赫克尔(Haeckel,1874)首次提出,后来又由梅契尼柯夫(1887)修正,海曼(Hyman,1940)又给以复兴。现分述如下:
1.赫克尔的原肠虫学说认为多细胞动物最早的祖先是由类似团藻的球形群体,一面内陷形成多细胞动物的祖先。这样的祖先,因为和原肠胚很相似,有两胚层和原口,所以赫克尔称之为原肠虫(gastraea)(图3-7)。
2.梅契尼柯夫的吞噬虫学说(实球虫或无腔胚虫学说)梅契尼柯夫观察了很多低等多细胞动物的胚胎发育,他发现一些较低等的种类,其原肠胚的形成主要不是由内陷的方法,而是由内移的方法形成的。同时他也观察了某些低等多细胞动物,发现它们主要是靠吞噬作用进行细胞内消化,很少为细胞外消化。由此推想最初出现的多细胞动物是进行细胞内消化,细胞外消化是后来才发展的。梅契尼柯夫提出了吞噬虫学说,他认为多细胞动物的祖先是由一层细胞构成的单细胞动物的群体,后来个别细胞摄取食物后进入群体之内形成内胚层,结果就形成为二胚层的动物,起初为实心的,后来才逐渐地形成消化腔,所以梅契尼柯夫便把这种假想的多细胞动物的祖先,叫做吞噬虫(phagocitella)(图3—7)。

这两种学说虽然在胚胎学上都有根据,但在最低等的多细胞动物中,多数是像梅契尼柯夫所说的由内移方法形成原肠胚,而赫克尔所说的内陷方法,很可能是以后才出现的。所以梅氏的学说容易被学者所接受。同时梅氏的说法看来更符合机能与结构统一的原则。不能想象先有一个现成的消化腔,而后才有进行消化的机能。可能是由于在发展过程中有了消化机能,同时逐渐发展出消化腔的。恩格斯说:“整个有机界在不断地证明形式和内容的同一或不可分离。形态学的现象和生理学的现象、形态和机能是互相制约的。”
从现有的原生动物看,其中鞭毛类动物形成群体的能力较强,如果原始的单细胞动物群体进一步分化,群体细胞严密分工协作,形成统一整体,这就发展成了多细胞动物。但是单细胞动物群体多种多样,有树枝状、扁平和球形的,前二者其个体在群体中的连接一般较疏松。根据多细胞动物早期胚胎发育的形状看,球形群体(类似团藻形状)与之一致,因此,群体学说认为由球形群体鞭毛虫发展成为多细胞动物符合于生物发生律。此外,从具鞭毛的精子普遍存在于后生动物,具鞭毛的体细胞在低等的后生动物间也常存在,特别是在海绵和腔肠动物,这些也可作为支持鞭毛虫是后生动物的祖先的证据。梅契尼柯夫所说的吞噬虫,很像腔肠动物的浮浪幼虫,它被称为浮浪幼虫样的祖先(planuloid ancestor)。低等后生动物是从这样一种自由游泳浮浪幼虫样的祖先发展的。根据这种学说,腔肠动物为原始辐射对称,可以推断它直接来源于浮浪幼虫样的祖先。扁虫两侧对称是后来发生的。
现在还有学者(Barnes,1987)认为,团藻样动物虽被作为鞭毛虫群体祖先的原型,但是这些具有似植物细胞的自养有机体不可能是后生动物的祖先,超微结构的证据表明,领鞭毛虫(cho- anoflagellates)原生动物更可能是后生动物的祖先。领鞭毛虫有些是单体的,有些是群体的。
最近10年有一个古老的学说在恢复生机,即Otto Butshli(1883)所提出的扁囊胚虫(plaku-la)学说(图3—7),他认为原始的后生动物是两侧对称的有两胚层的扁的动物,称此动物为扁囊胚虫。根据Butshli的看法,扁囊胚虫通过腹面细胞层的蠕动、爬行、摄食,最后该动物背腹细胞层分开成为中空的,这样逐渐地腹面的营养细胞内陷形成消化腔,同时产生了内外胚层,形成了两胚层动物。这里所提的扁囊胚虫与现存的扁盘动物丝盘虫(trichoplax)是相似的。有些学者认为丝盘虫是扁囊胚虫现存种类的证据。

(二)合胞体学说(Syncytial theory)这一学说主要是由Hadzi(1953)和Hanson(1977)提出的,认为多细胞动物来源于多核纤毛虫的原始类群(图3—8)。后生动物的祖先开始是合胞体结构,即多核的细胞,后来每个核获得一部分细胞质和细胞膜形成了多细胞结构。由于有些纤毛虫倾向于两侧对称,所以合胞体学说主张后生动物的祖先是两侧对称的,并由其发展为无肠类扁虫,认为无肠类扁虫是现在生存的最原始的后生动物。对该学说,持反对意见者较多,因为任何动物类群的胚胎发育都未出现过多核体分化成多细胞的现象,实际上无肠类合胞体是在典型的胚胎细胞分裂之后出现的次生现象。最主要的反对意见是不同意将无肠类扁虫视为最原始的后生动物。体型的进化是从辐射对称到两侧对称,如果认为无肠类扁虫两侧对称是原始的,那么腔肠动物的辐射对称倒成为次生的,这显然与已揭明的进化过程是相违背的。
此外,还有共生学说(symbiosis theory)(图3—8),认为不同种的原生生物共生在一起,发展成为多细胞动物。这一学说存在一系列的遗传学问题,因为不同遗传基础的单细胞生物如何聚在一起形成能繁殖的多细胞动物,这在遗传学上是难以解释的。
对多细胞动物起源,多数进化理论者倾向于单元说,但事实上已有一些提示,认为多细胞动物的来源是多元的。即起源于不止一类原生动物的祖先。这些观点的大部分集中在是鞭毛虫还是纤毛虫是祖先类群。并仍在找寻从原生动物过渡到多细胞动物的中间类型。
复习题
1.一般了解中生动物的简要特征以及对其分类地位的不同看法。
2.根据什么说多细胞动物起源于单细胞动物?
3.初步掌握多细胞动物胚胎发育的共同特征(从受精卵、卵裂、囊胚、原肠胚、中胚层与体腔形成、胚层分化等方面)。
4.什么叫生物发生律?它对了解动物的演化与亲缘关系有何意义?
5.关于多细胞动物起源有几种学说?各学说的主要内容是什么?哪个学说易被多数人接受,为什么?你的看法如何?
本文标题:第三章 多细胞动物的起源
手机页面:http://m.dljs.net/dlsk/dongwuxue/59430.html
本文地址:http://www.dljs.net/dlsk/dongwuxue/59430.html
