第一节 腔肠动物门的主要特征
多孔动物在动物演化上是一个侧支;腔肠动物才是真正后生动物的开始。这类动物在进化过程中占有重要位置,所有其他后生动物都是经过这个阶段发展起来的。腔肠动物为辐射对称、具两胚层、有组织分化、原始的消化腔及原始神经系统的低等后生动物(metazoa)。
(一)辐射对称 多孔动物的体型多数是不对称的。从腔肠动物开始,体型有了固定的对称形式。本门动物一般为辐射对称(radial symmetry)。即大多数腔肠动物,通过其体内的中央轴(从口面到反口面)有许多个切面可以把身体分为2个相等的部分。这是一种原始的低级的对称形式。这种对称只有上、下之分,没有前后左右之分,只适应于在水中营固着的或漂浮的生活。利用其辐射对称的器官从周围环境中摄取食物或感受刺激。在腔肠动物中有些种类已由辐射对称发展为两辐射对称(biradial symmetry),即通过身体的中央轴,只有两个切面可以把身体分为相等的两部分。这是介于辐射对称和两侧对称的一种中间形式。
(二)两胚层、原始消化腔 多孔动物虽然具有二胚层,但从发生来看,它与其他后生动物不同,因此一般只称为二层细胞。腔肠动物才是具有真正二胚层(内、外胚层)的动物。在二胚层之间有由内、外胚层细胞分泌的中胶层。由内外胚层细胞所围成的体内的腔,即胚胎发育中的原肠腔。它与海绵的中央腔不同,具有消化的功能,可以行细胞外及细胞内消化。因此,可以说从这类动物开始有了消化腔。这种消化腔又兼有循环的作用,它能将消化后的营养物质输送到身体各部分,所以又称为消化循环腔(gastrovascular cavity)。有口、没有肛门,消化后的残渣仍由口排出。它的口有摄食和排遣的功能。口即为胚胎发育时的原口,与高等动物比较,可以说腔肠动物相当于处在原肠胚阶段。
(三)组织分化 海绵动物主要是有细胞分化。腔肠动物不仅有细胞分化,而且开始分化出简单的组织。动物的组织一般分为上皮、结缔、肌肉、神经四类,而在腔肠动物上皮组织却占优势,由它形成体内、外表面,并分化为感觉细胞、消化细胞等。它的特点是在上皮细胞内包含有肌原纤维。这种细胞具有上皮和肌肉的功能,所以称为上皮肌肉细胞(epithelio-muscular cell)简称皮肌细胞。同时腔肠动物的上皮还具有像神经一样的传导功能,这是近些年来应用电生理学技术和电子显微镜检来研究腔肠动物神经的一个发现。非神经的传导(non-nervous conduc-tion)或类神经(neuroid)传导,首先是在腔肠动物得到证实的。
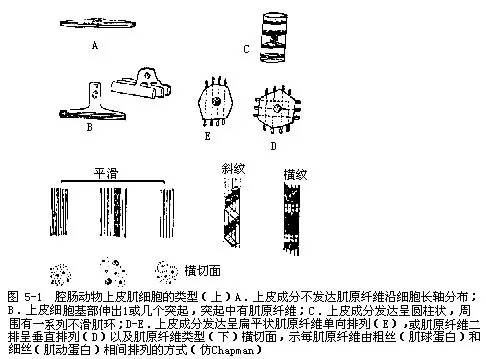
(四)肌肉的结构(musculature) 上皮肌肉细胞既属于上皮,也属于肌肉的范围。这表明上皮与肌肉没有分开,是一种原始的现象。一般在上皮肌肉细胞的基部延伸出一个或几个细长的突起,其中有肌原纤维(myofibrils)(图5-1B),也有的上皮成分不发达,成为肌细胞(myocyte)(图5-1A),有的是上皮成分发达,细胞呈扁平状,肌原纤维呈单向排列(图5-1E),或者是2排肌原纤维呈垂直排列(图5-1D),也有的上皮成分发达呈圆柱状,周围有一系列的平滑肌环(图5-1C)。肌纤维也分为横纹肌、斜纹肌和平滑肌(图5-1)。每个肌原纤维都是由一束细丝组成,这些丝又分粗、细2种,与高等动物粗(肌球蛋白)、细(肌动蛋白)丝相似,其收缩机理也和高等动物的相似。关于肌肉的神经支配了解的不多,近年来有的实验证明,腔肠动物的神经与肌肉的接触部分——神经肌肉突触(neuromuscular synapses)的超微结构和神经肌肉连接(neuromus-cular junction),也都与高等动物的相似。
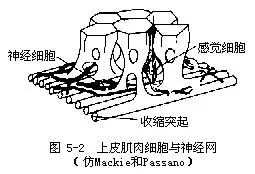
(五)原始的神经系统——神经网(nerve net) 是动物界里最简单最原始的神经系统。一般认为它基本上是由二极和多极的神经细胞组成。这些细胞具有形态上相似的突起,相互连接形成一个疏松的网,因此称神经网。有些种类只有一个神经网,存在于外胚层的基部(图5-2);有些种类有2个神经网,分别存在于内、外胚层的基部;还有些除了内外胚层的神经网外,在中胶层中也有神经网(图5-3)。神经细胞之间的连接,经电子显微镜证明,一般是以突触相连接,也有非突触的连接。这些神经细胞又与内、外胚层的感觉细胞、皮肌细胞等相连系。感觉细胞接受刺激,通过神经细胞传导,皮肌细胞的肌纤维收缩产生动作,这种结合形成神经肌肉体系(neuro-muscular system)。这样对外界刺激(光、热、化学的、机械的、食物等)产生有效的反应,如捕食、避敌以及协调整体的活动等。但腔肠动物没有神经中枢,神经的传导一般是无定向的,因此称为扩散神经系统(diffuse nervous system)。同时,神经的传导速度也较慢,它比人的神经传导速度约慢1000倍以上,这都说明这种神经系统的原始性。
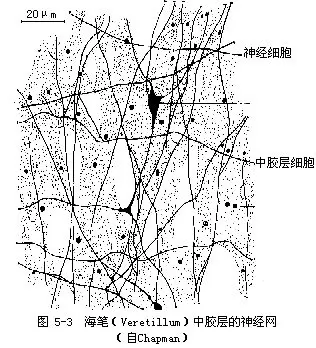
近年来对腔肠动物神经突起的超微结构的研究,看到神经连接的突触,在形态上有极化现象,就是只在神经交接的一个突起上有泡,而另一个没有。在没有极化的突触上,2个突起都有泡。这种形态上的极化可能是传导系统中极化传导的基础。
第二节 腔肠动物门代表动物——水螅(Hydra)
水螅生活在淡水中,在水流较缓水草丰富的清水中常可采到。水螅分布较广、容易采集和培养,且便于观察其结构,因此常用作实验材料。通过它可了解这类动物的基本结构。
水螅体为圆柱状,能伸缩,遇到刺激时可将身体缩成一团。一端附于水草或其他物体上,附着端称为基盘(basal or pedal disk)。另一端有口,口长在圆锥形的突起——垂唇(hypostome)上,平常口关闭呈星形,当摄食时口张开,在口之周围,有细长的触手(tentacle),一般6~10条,呈辐射排列,主要为捕食器官(图5-4)。当水螅饥饿时,触手伸得很长。如狩猎一样,捕到食物后由触手缩回来送到口中。也可借助于触手和身体弯曲作尺蠖样运动或翻筋斗运动(图5-5)。

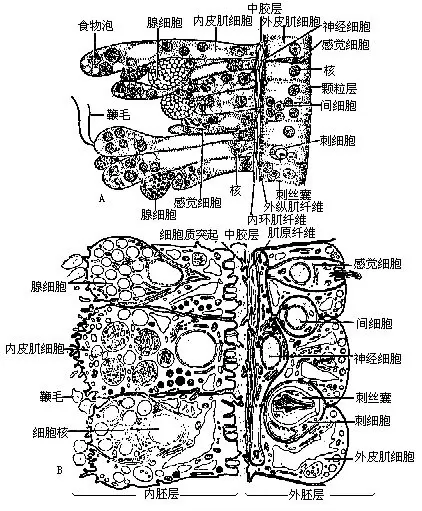
水螅的身体内部为一空腔,由口与外界相通,也与触手相通,此为消化循环腔。其体壁由两层细胞构成,在2层细胞之间为中胶层。体表的一层为外胚层,这层细胞主要有保护和感觉的功能。里面的一层为内胚层,主要有营养功能(图5-6)。

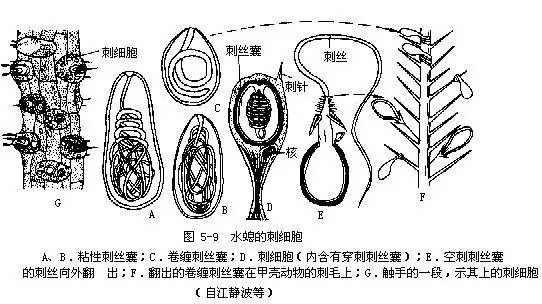
外胚层包括皮肌细胞(称上皮肌细胞或外皮肌细胞)、腺细胞、感觉细胞、神经细胞、刺细胞和间细胞(图5-7)。在外胚层中皮肌细胞数目最多,皮肌细胞基部的肌原纤维沿着身体之长轴排列,如一层纵行的肌纤维,它收缩时可使水螅身体或触手变短。感觉细胞(sensory cell)分散在皮肌细胞之间,特别在口周围、触手和基盘上较多,其体积很小,细胞质浓,端部有感觉毛,基部与神经纤维连接。神经细胞(nerve cell)位于外胚层细胞的基部,接近于中胶层的部分,神经细胞的突起彼此连接起来形成网状(图5-8),传导刺激向四周扩散,所以当其身体的一部分受较强的刺激时,全身都发生收缩反应,以避开有害刺激。刺细胞(cnidoblast)是腔肠动物所特有的,它遍布于体表,触手上特别多。每个刺细胞有一核位于细胞之一侧,并有囊状的刺丝囊(nematocyst),囊内贮有毒液及一盘旋的丝状管。水螅有四种刺丝囊(图5-9):穿刺刺丝囊,其中有一条细长中空的刺丝,当受刺激时,刺丝向外翻出,像手套的指端从内向外翻一样,就可把毒素射入捕获物或其他小动物体内,将其麻醉或杀死。卷缠刺丝囊,不注射毒液,而只缠绕被捕物。还有2种粘性刺丝囊,对捕食和运动有作用。间细胞(interstitial cell),主要在外胚层细胞之间,有一堆堆的小细胞,大小与皮肌细胞的核差不多,一般认为它是一种未分化的胚胎性的细胞,可以分化成刺细胞和生殖细胞等。腺细胞(gland cell)身体各部都有,以基盘和口周围最多,能分泌粘液,可使水螅附着于物体上或在其上滑行。也可分泌气体,由粘液裹成一气泡,使水螅由水底上升至水面。

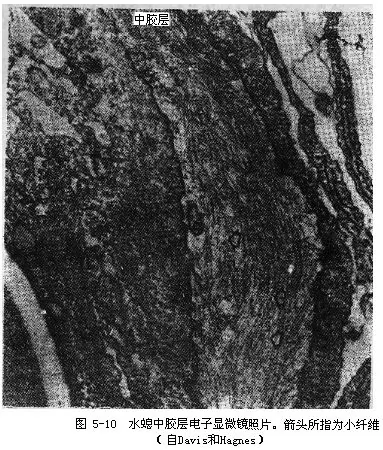
中胶层薄而透明,为内外胚层细胞分泌的胶状物质,在身体和触手都是连续的。在电子显微镜下,中胶层中有很多小纤维,皮肌细胞突起也伸入其中(图5-10)。中胶层像是有弹性的骨骼,对身体起支持作用。
内胚层包括内皮肌细胞、腺细胞和少数感觉细胞与间细胞(图5-7)。在内胚层细胞的基部也有分散的神经细胞,但未连接成网。内皮肌细胞或称营养肌肉细胞(nutritivemus cular cell),是一种具营养机能兼收缩机能的细胞,在细胞之顶端通常有2条鞭毛(1~5条),由于鞭毛的摆动能激动水流,同时也可伸出伪足吞食食物,细胞内常常有不少食物泡,其基部的肌原纤维,沿着体轴或触手之中心呈环形排列,收缩时可以使身体或触手变细。在口周围,皮肌细胞的肌原纤维还有括约肌的作用。腺细胞在内皮肌细胞之间,分散于内胚层各部分。腺细胞所处的部位不同,其功能也不一样,如在垂唇部分的可分泌粘液,在滑润作用,使食物容易被吞进去;在消化循环腔内的,则能分泌消化酶消化食物。
水螅以各种小甲壳动物(如溞类、剑水蚤等)、小昆虫幼虫和小环节动物等为食。以触手捕食,被捕的食物可比水螅大很多倍。触手将捕获物移向口部,由于捕获物受刺丝的损伤放出谷胱甘肽(glutathione),在该物质的刺激下,水螅口张开,食物进入消化循环腔。如把商品谷胱甘肽放在盛有水螅的水中,即使没有捕获物,水螅也表现出取食的动作。在消化腔内,由腺细胞分泌酶(主要为胰蛋白酶)进行细胞外消化(extracellular digestion),经消化后形成一些食物颗粒,由内皮肌细胞吞入进行细胞内消化。食物大部分在细胞内消化。消化后的食物可储存在内胚层细胞或扩散到其他细胞。不能消化的残渣再经口排出体外。
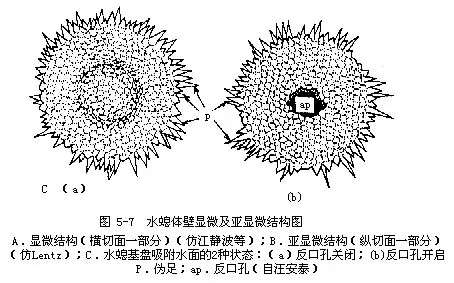
多年来一般认为腔肠动物(包括水螅类)只通过口与外界相通。但有少数学者发现水螅属(Hydra)、柄水螅属(Pelmatohydra)和绿水螅属(Chlorahydra)的种类,在基盘中央有一反口孔(aboral pore)。观察水螅基盘的超微结构时发现,肌原纤维以反口孔为中心呈定向辐射状排列。这种排列方式有利于控制反口孔的开启和关闭。反口孔静止时,基盘附着层外表中心看不到孔迹(图5-7C),反口孔开启时,有废物或气体从孔内排出。看来水螅的反口孔具有肛门的部分生理功能。
呼吸和排泄,没有特殊的器官,由各细胞吸氧、排出二氧化碳和废物。
水螅的生殖分无性和有性两种。经常进行无性生殖——出芽生殖。即体壁向外突出,逐渐长大,形成芽体。芽体的消化循环腔与母体相通连,芽体长出垂唇、口和触手,最后基部收缩与母体脱离,附于他处营独立生活。有性生殖是精卵结合。大多数种类为雌雄异体,少数为雌雄同体。生殖腺是由外胚层的间细胞分化形成的临时性结构,精巢为圆锥形,卵巢为卵圆形(图5-11)。卵巢内一般每次成熟一个卵,也有的种类一次成熟几个卵。卵在成熟时,卵巢破裂,使卵露出。精巢内形成很多精子,成熟的精子,出精巢后,游近卵子与之受精。受精卵进行完全卵裂,以分层法形成实心原肠胚。围绕胚胎分泌一壳,从母体上脱落下来,沉入水底,渡过严冬或干旱等条件,至春季或环境好转时,胚胎完成其发育。壳破裂,胚胎逸出,发育成小水螅(图5-12)。


腔肠动物的再生能力很强,如把水螅切成几小段,每段都能长成一个小水螅,但是只有单独的触手不能再生成完整的动物。通过水螅的垂唇和口切开,能长成双头水螅(图5-12)。近年来有人将水螅(Hydra oligactis,H.pseudoligactis)内外胚层细胞分开,结果2层细胞都各自再生成为完整的水螅。在水螅的再生和出芽方面,过去认为间细胞起重要的不可缺少的作用,由间细胞分化为各种类型的细胞。现在有人认为,间细胞在再生和出芽作用中不是不可缺少的,如用X射线或含氮芥子气(nitrogen mustard)处理破坏间细胞,仍能进行再生或出芽。

第三节 腔肠动物门的分纲
腔肠动物约10000多种。分为3纲:水螅纲、钵水母纲、珊瑚纲。
一、水螅纲(Hydrozoa)
本纲动物绝大多数生活在海水中,少数生活在淡水。生活史中大部分有水螅型和水母型,即有世代交替现象。如薮枝虫(Obelia)(图5-14)。

薮枝虫生活于浅海,固着在海藻、岩石或其他物体上,为一树枝状的水螅型群体(图5-13)。群体基部的构造很像植物的根,故称螅根(hydrorhiza),由螅体上生出很多直立的茎,称为螅茎(hydrocaulus)。螅茎上分出2种个体——水螅体(hydranth)与生殖体(gonangium)。整个群体外面,包围着由外胚层分泌的一层透明的角质膜,称围鞘(perisarc),具保护和支持的功能。水螅体主要管营养,其构造与水螅基本相同,有口及触手,触手是实心的,垂唇较水螅的长大,其外有一透明的杯形鞘,称为水螅鞘(hydrotheca)。生殖体无口及触手,只有一中空的轴,称为子茎(blas-tostylus),子茎的周围,有透明的瓶状鞘,称为生殖鞘(gonotheca)。生殖体能行无性生殖,其营养主要靠水螅体供给,因为水螅体和生殖体彼此由螅茎中的共肉(coenosarc)连接,整个群体的消化循环腔是相通连的。群体中任一水螅体捕食消化后,可通过消化循环腔输送给其它部分或其他个体。
生殖体成熟后,子茎以出芽的方法产生许多水母芽,水母芽成熟,脱离子茎,由生殖鞘顶端的开口出来,在海水中营自由生活,其结构较简单,很小,约1mm~2mm,体形如一圆伞,伞边缘生有很多细的触手(初生时16个),下伞面中央有一短的垂唇,口向内通到胃,再由胃伸出4个辐管,与伞边缘的环管相通。口、胃、辐管、环管构成水母的消化循环系。水螅水母特征之一是在伞下面边缘有一圈薄膜称为缘膜(velum),而薮枝虫的水母缘膜退化。在伞边缘有8个平衡囊,司平衡。在4条辐管上有四个由外胚层形成的精巢或卵巢(雌雄异体)。精卵成熟后在海水中受精。受精卵发育,以内移的方式形成实心的原肠胚,在其表面生有纤毛,能游动,称为浮浪幼虫(planula)。浮浪幼虫游动一段时期后,固着下来,以出芽的方式发育成水螅型的群体。
薮枝虫的生活史经过2个阶段。水螅型群体以无性出芽的方法产生单体的水母型,水母型又以有性生殖方法产生水螅型群体,这两个阶段互相交替,完成世代交替的生活史。
由薮枝虫的结构与生活史可看出本纲动物的主要特征:
1.一般是小形的水螅型或水母型动物。
2.水螅型结构较简单,只有简单的消化循环腔。
3.水母型有缘膜,触手基部有平衡囊。
4.生活史大部分有水螅型与水母型,即有世代交替现象(如薮枝虫),少数种类水螅型发达,无水母型(如水螅)或水母型不发达(如筒螅Tubularia),也有水母型发达,水螅型不发达或不存在,如钩手水母(Gonionemus)、桃花水母(Craspedacusta);还有的群体发展为多态现象,如僧帽水母(Physalia)(图5-14,15)。

二、钵水母纲(Scyphozoa)
本纲动物全部生活在海水中,大多为大型的水母类(如有一种霞水母 Cyanea arctica伞部直径大的有2m多,触手长30多m),水母型发达,水螅型非常退化,常常以幼虫的形式出现,而且水母型的构造比水螅水母复杂。如海月水母。
海月水母(Aurelia aurita Lamarck)(图5-17)营漂浮生活,体为白色透明的盘状,在伞的边缘生有触手,并有8个缺刻,每个缺刻中有一个感觉器,也称触手囊,囊内有钙质的平衡石(stato-lith),囊上面有眼点(ocellus),囊下面有缘瓣(lappet),缘瓣上有感觉细胞和纤毛,另外有2个嗅窝(图5-18)。当水母体不平衡时,触手囊对感觉纤毛的压力不同,而产生不平衡的感觉。在内伞的中央有一呈四角形的口,由口的四角上伸出 4条口腕(oral lobe)。消化循环系统比较复杂,由口进去为胃腔,位于体中央,向四方扩大成四个胃囊,由胃囊上和胃囊之间伸出分枝的和不分枝的辐管(radial canal),这些辐管均与伞边缘的环管(ring canal)相连。水流由口进去至胃腔,经过一定的辐管至环管,然后再由一定的辐管流至胃囊,经口流出。在胃囊的里面,有四个由内胚层产生的马蹄形的生殖腺,位于胃囊底部的边缘,在生殖腺内侧,长有很多丝状的结构称为胃丝,也是由内胚层形成的,其上有很多刺细胞,食物进入胃囊后,即被刺丝杀死,经消化后(细胞内和细胞外消化)由辐管分布到全身各部,胃丝也起着保护生殖腺的作用。


由生殖腺产生精子或卵(钵水母为雌雄异体),精子成熟后随水流至雌体内受精,也有的在海水中受精,受精卵经完全均等卵裂形成囊胚,再由内陷方式形成原肠胚,此时胚胎表面长出纤毛,成为浮浪幼虫,在海水中游动一个时期后,附于海藻或其他物体上,发育成小的螅状幼体(hydru-la),有口和触手,可营独立生活,然后进行横裂,由顶而下分层为钵口幼体(scyphistoma),再进行连续横分裂形成一个个碟状个体,称横裂体(strobila)。横裂体成熟后一个个依次脱落下来,称为碟状幼体(ephyra),由它发育成水母成体。由此可见,钵水母的生活史(图5-19)虽有世代交替,但水母型发达而水螅型则退化成为幼虫。也有的钵水母无世代交替现象,只有水母型。
钵水母与水螅水母的主要不同点在于:
1.钵水母一般为大形水母,而水螅水母为小形的。
2.钵水母无缘膜,而水螅水母有缘膜。钵水母的感觉器官为触手囊,水螅水母为平衡囊。
3.钵水母的结构较复杂,在胃囊内有胃丝,而水螅水母则无。

4.钵水母的生殖腺来源于内胚层,水螅水母的生殖腺来源于外胚层。
钵水母类在腔肠动物中是经济价值较高的一类动物,比如海蜇即属此类。海蜇(Rhopilemaesculentum)(图5-20)的结构与海月水母基本上是一致的,不同的是:海蜇的伞为半球形,中胶层很厚,含有大量的水分和胶质物,伞的边缘无触手,由8个感觉器将伞缘平分为8个区,每个区的伞缘有14~20个小舌状的缘瓣(lappets,其数目为种的分类依据之一)。口腕愈合,大形口消失,在口柄的基部有8对翼状肩板,在口柄下部各有8个口腕,每个口腕又分成三翼,在其边缘上形成很多小孔,称为吸口。在口腕上长有很多触手(或称丝状附属器、棒状附属器),棒状附属器与腕管相通。各腕末端有一条长的棒状附属器。肩板上也有很多吸口及触手。海蜇就是靠吸口吸食一些微小的动植物为食物,由吸口周围的触手先把小动物麻痹或杀死,然后送入口,经口腕中很多分枝的小管到胃腔,这种构造和取食的情况有点像植物根吸收养料,因此将海蜇这一类的钵水母归为一目,称为根口水母目(Rhizostomae)。
关于海蜇的生活史(图5-21),近年来我国已作了详细的报道。这对繁殖生物学和发育生物学的研究,对海蜇资源的预测预报和增殖都有重要意义。
海蜇的营养价值较丰富,含有蛋白质、维生素B1、维生素B2等。经加工处理后的蜇皮,是海蜇的伞部,蜇头或蜇爪为海蜇的口柄部分。我国食用海蜇的历史悠久,在我国沿海海蜇的产量非常丰富,浙江、福建沿海一带最多。


除海蜇外,大多数的钵水母对渔业生产有害,不仅危害幼鱼、贝类,且能破坏网具。如 1952年在福建连江沿海发生了一种钵水母为灾的情况。渔民的网具被破坏后几月不能生产。腔肠动物的刺丝囊对人的危害很大,如一些大的水母或海蜇螫刺人体后,可造成严重创伤。对刺丝囊里的毒性物质的成分性质,近年来已进行了一些研究。目前正在研究很多海产种类的刺丝囊毒素,作为新的药物来源或其他生物医学化合物。从1972年就已发现在4种腔肠动物的提取物中有抗肿瘤的药物。


仿生学也在研究水母,制作预测风暴的报警仪器。以前预测海上风暴要用雷达站、水声站,甚至气象卫星进行综合观测,十分不便,而生活在海水中的水母在进化过程中发展了一套预测风暴的报警装置,使它在风暴来临前许多小时就游向大海。模仿水母感觉器的风暴预报仪器能提前15个小时作出预报,并指出风暴来的方向,装置简单,操作方便。又如海蜇的运动是由脉冲式的喷射而推进的,而喷气式飞机是连续不断的气流喷射而推进的。有的科学家曾设想把海蜇的推进方式用于喷气式飞机的设计,这样既能节省能量,又能最好的利用所产生的动力。

三、珊瑚纲(Anthozoa)
这纲动物与前二纲不同,只有水螅型,没有水母型,且水螅体的构造较水螅纲的螅体复杂。全为海产,多生活在暖海、浅海的海底。构成“海底花园”的主要为珊瑚虫。一般所见到的珊瑚为其骨骼。沿海常见的为海葵。
海葵(图5-22)无骨骼,身体呈圆柱状,一端附于海中岩石或其他物体上,该端称为基盘。另一端有口,呈裂缝形,口周围部分称为口盘,其周围有几圈触手,触手上有刺细胞,可用以捕食鱼虾及活的小动物。捕捉食物后经口,进入口道(stomodaeum),口道壁是口部的外胚层细胞褶入形成的(此为进化现象)。在口道的两端各有一纤毛沟( siphonoglyphe)或称口道沟(有些种类只有一个纤毛沟),纤毛沟内壁的细胞具纤毛。当海葵收缩成一团时,水流仍可由纤毛沟流入消化循环腔。
消化循环腔的构造较复杂,其中有宽、窄不同的隔膜(mesentery),隔成很多小室。隔膜是由体壁上内胚层细胞增多向内突出形成的,在2层内胚层之间为中胶层。隔膜的作用,主要为支持,并增加消化面积。根据隔膜的宽度可分一、二、三级,只有一级隔膜与口道相连。在隔膜游离的边缘有隔膜丝(mesenteric filament),隔膜丝沿隔膜的边缘下行,一直达到消化循环腔的底部;有的达底部时形成游离的线状物称为毒丝,其中含有丰富的刺细胞,当动物收缩时经常由口或壁孔射出,有防御及进攻的机能(图5-23)。隔膜丝主要由刺细胞和腺细胞构成,能杀死摄入体内的捕获物,并由腺细胞分泌消化液,行细胞外消化和细胞内消化。肌肉较发达,在较大的隔膜上都有一纵肌肉带称为肌旗(muscle band),隔膜和肌旗的排列是分类的依据之一。

海葵为雌雄异体,生殖腺长在隔膜上接近隔膜丝的部分,由内胚层形成。精子成熟后,由口流出,进入另一雌体内与卵结合形成受精卵,也有的在海水中受精,在母体内发育形成浮浪幼虫,出母体,游动一时期后,固着下来发育成新个体。但也有的海葵不经浮浪幼虫,直接发育成为海葵后出母体。无水母型。无性生殖为纵分裂或出芽。
从海葵可了解珊瑚虫的基本结构,它与水螅纲的螅型体的不同点在于:
1.珊瑚纲只有水螅型,其构造较复杂,有口道、口道沟、隔膜和隔膜丝。水螅纲的螅型体构造较简单,只有垂唇,无上述结构。
2.珊瑚纲螅型体的生殖腺来自内胚层,水螅纲螅型体的生殖腺来自外胚层。
海葵是单体的,无骨骼。很多珊瑚虫为群体,大多具骨骼。
珊瑚骨骼的形成大多数珊瑚虫的外胚层细胞能分泌骨骼。在八放珊瑚亚纲(Octocorallia)(触手和隔膜各8个),由外胚层的细胞移入中胶层中分泌角质或石灰质的骨针或骨片。这些骨针存在于中胶层中或突出于体表面,如海鸡冠(Alcyonium)和海鳃(Pennatula)。有的种类小骨片连接成管状的骨骼,如笙珊瑚(Tubipora)。还有的骨针或骨片愈合成中轴骨,如红珊瑚(Cor-allium)(图5-23,图5-24)。常见的六放珊瑚亚纲(Hexacorallia)(触手和隔膜一般为6的倍数)的石珊瑚目(Medreporaria)有单体与群体,每个虫体与海葵相似,其基盘部分与体壁的外胚层细胞能分泌石灰质物质,积存在虫体的底面、侧面及隔膜间等处,好像每个虫体都坐在一个石灰座上,称为珊瑚座(corallite)(图5-25),如石芝(Fungia)。群体珊瑚虫其共肉部分的外胚层也分泌石灰质,由于群体的形状不同,其骨骼的形状也不一样。有的为树枝状,如鹿角珊瑚(Madrepora=Acropora),有的为圆块状,如脑珊瑚(Meandrina)(图 5-26)。


石珊瑚的骨骼是构成珊瑚礁和珊瑚岛的主要成分。由大量的珊瑚骨骼堆积成的岛屿,如我国的西沙群岛、印度洋的马尔代夫岛、南太平洋的斐济群岛等。造礁的石珊瑚虫内胚层细胞中常有大量的单细胞虫黄藻(Zooxanthella)与其共生,有利于珊瑚虫从虫黄藻补充氧气和糖类,加速骨骼的生长(虫黄藻获得珊瑚虫的代谢废物如CO2、氮等,以利其进行光合作用)。石珊瑚的生活习性要求温暖(一般要求水温22℃~30℃)、浅水(水深约在45m以内)的环境,海水对它有一定的冲击力量,靠海边的珊瑚承受海水冲击的部分生活得最好,所以随着骨骼的堆积,常沿着海岸逐渐向海里推移,逐渐扩展,形成大的岛屿。在沿海的岸礁,有如海边上的天然长堤,能使海岸坚固。但在海底的暗礁,可妨碍航行。


石珊瑚还可用来盖房子,如海南沿海一带用珊瑚建造的房子坚固耐用、便宜美观。还可用石珊瑚烧石灰制水泥、铺路等。我国台湾的许多街道都是用石珊瑚铺成的,路面坚固平坦。还用来养殖石花菜,或作观赏用、制作装饰品等,总之这类珊瑚的用途是很广的。
珊瑚骨骼对地壳形成也有一定作用。在地质上常见到石灰质珊瑚骨骼形成的石灰岩,一般称为珊瑚石灰岩。有这种石灰岩的地方,说明这里在亿万年以前曾经是温暖的浅海。如我国四川、陕西交界的强宁、广元间就有这种石灰岩,考证其地质年代应在志留纪。古珊瑚礁和现代珊瑚礁可形成储油层,对找寻石油也有重要意义。
第四节 腔肠动物的系统发展

腔肠动物是真正多细胞动物的开始。从其个体发育看,一般海产的腔肠动物,都经过浮浪幼虫的阶段,由此可推想:最原始的腔肠动物是能够自由游泳的、具纤毛的动物,其形状像浮浪幼虫,即梅契尼柯夫所假设的群体鞭毛虫,细胞移入后形成为原始二胚层的动物,发展成腔肠动物。

在现存的腔肠动物中,水螅纲无疑是最低等的一类,因为其水螅型与水母型的构造都比较简单,生殖腺来自外胚层。钵水母纲水螅型退化,水母型发达,结构较复杂。珊瑚纲无水母型,只有结构复杂的水螅型。后2纲的生殖腺又都来自内胚层,因此可以认为,钵水母纲和珊瑚纲可能起源于水螅纲,沿着不同的途径发展而来的。
综上所述,腔肠动物门的内容可简要概括如下:
腔肠动物一般为辐射对称(也有两侧辐射对称的),具两胚层,有原始的消化腔(消化循环腔),有口无肛门,行细胞外及细胞内消化。有组织分化,具原始的肌肉结构(皮肌细胞)和原始的神经系统(神经网),有刺细胞。有骨骼时,为钙质或角质。体型一般为水螅型和水母型,水螅型适于固着生活,体呈圆筒形,水母型适于漂浮生活,体一般呈盘形,二者的结构基本相同(图5-27),如把水螅型倒置,其基部与水母的外伞相当,沿中轴压平即成盘形。二者之不同点:水母型适应漂浮生活,中胶层加厚,这可以减轻身体的比重。也有些为多态的群体。在生活史中,有些具发达的水螅型与水母型,有世代交替现象;有些水母型发达,水螅型不发达或不存在;有些水螅型发达,水母型不发达或不存在。海产种类个体发育中经浮浪幼虫时期。过去一般认为有些种类有经济价值,随着科学的发展,发现腔肠动物还有多方面的用途,值得探索。
附:栉水母动物门(Ctenophora)
栉水母过去曾列入腔肠动物门,作为无刺胞亚门,或作为栉水母纲。现在一般把它另列为一门。也有人把它与腔肠动物门并列,都称为辐射动物。
这一类动物的种类(不到100种)和数量都比较少,全部生活在海水中,能发光,营浮游生活,也有的能爬行。体形有球形、瓜形、卵圆形以及扁平带状等(图5-28)。形态结构像腔肠动物的有:体型基本上属于辐射对称,但两侧辐射对称很明显;身体也分内、外胚层及中胶层;消化循环腔与钵水母的相似,具有分枝的辐管,除此之外,体内没有其他腔。

本门动物也具有其他动物所没有的特点:
1.栉水母体表具有8行纵行的栉板(combplate)(图5-29),每一栉板是由一列基部相连的纤毛所组成。栉板下面有肌纤维使栉板运动,栉板为运动器。

2.有触手的栉水母在体两侧各有一触手囊或称触手鞘(tentacle sheath),囊内各有一条触手,触手上没有刺细胞(Euchlora rubra例外,有刺细胞,无粘细胞),而有大量的粘细胞(colloblast),该细胞表面分泌粘性物质,可用以捕食,细胞内侧有螺旋状丝,捕获物被粘着后,不致因其挣扎而损坏细胞(图5-30)。

3.在反口面有一集中的感觉器官(sense organ),结构较复杂(图5-31)。在平衡囊内,由4条平衡纤毛束支持一个钙质的平衡石(statolith),在平衡纤毛束基部有纤毛沟(ciliated furrow)和8行纵行的栉板相连。栉水母的感觉器官是司平衡的器官。

4.神经系统也较集中,虽然在外胚层基部有神经网,但已向8行栉板集中,形成8条辐射神经索。
5.胚胎发育中,可认为已开始出现不发达的中胚层细胞,由它发展成肌纤维。
由以上看出,栉水母类在进化上为特殊的一群,与腔肠动物接近,但较腔肠动物略为高等。有的学者认为爬行栉水母可能进化为扁形动物,但一般认为栉水母类在进化上是一盲端支流,与高等动物没有直接关系。
栉水母类以浮游生物为食,同时它本身又是鱼类的饵料,因此它们在食物链中有一定作用。栉水母类还能吃牡蛎幼虫、鱼卵和鱼苗,对牡蛎养殖和某些鱼的繁殖有一定影响。
这类动物虽然数量不多,又是进化的盲端,但却引起了科学家们对它的兴趣,希望用它来解决一些一般生物学问题,并已取得一定的成果。比如近年来发现栉水母有4种传导系统(栉板活动系统、上皮下神经网的栉板抑制系统、肌细胞间的传导系统、协调发光的系统),希望以此来解决由原始的传导系统如何产生出这4种传导系统,和以后在神经传导进化过程中又如何产生出像高等动物那样一个中枢神经系统。可见在这个很特化的小类群,相当简单的协调系统却显示出了行为控制和进化的一般原理。
复习题
1.腔肠动物门的主要特征是什么?如何理解它在动物进化上占重要位置?
2.掌握水螅的基本结构如内外胚层细胞的分化等,通过它了解腔肠动物的体壁结构、组织分化等基本特征。
3.腔肠动物分哪几个纲,各纲的主要特征是什么?有何价值?
4.初步了解腔肠动物的系统发展。
本文标题:第五章 腔肠动物门(Coelenterata)
手机页面:http://m.dljs.net/dlsk/dongwuxue/59428.html
本文地址:http://www.dljs.net/dlsk/dongwuxue/59428.html
