第一节 软体动物门的主要特征
软体动物的形态结构变异较大,但基本结构是相同的。身体柔软,不分节,可区分为头、足、内脏团3部分,体外被套膜,常常分泌有贝壳。
(一)身体的划分软体动物的身体一般可分为头、足和内脏团3部分(图9-1)
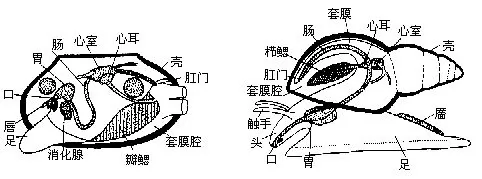
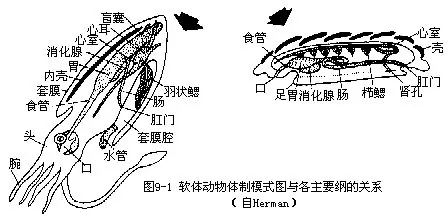
1.头部 位身体的前端。运动敏捷的种类,头部分化明显,其上生有眼、触角等感觉器官,如田螺、蜗牛及乌贼等;行动迟缓的种类,头部不发达,如石鳖;穴居或固着生活的种类,头部已消失,如蚌类、牡蛎等。
2.足部 通常位于身体的腹侧,为运动器官,常因动物的生活方式不同,而形态各异。有的足部发达,呈叶状、斧状或柱状,可爬行或掘泥沙;有的足部退化,失去了运动功能,如扇贝等;固着生活的种类,则无足,如牡蛎;有的足已特化成腕,生于头部,为捕食器官,如乌贼和章鱼等,称为头足;少数种类足的侧部(即侧足parapodium)特化成片状,可游泳,称为翼或鳍,如翼足类(Pteropoda)。
3.内脏团(visceral mass)为内脏器官所在部分,常位于足的背侧。多数种类的内脏团为左右对称,但有的扭曲成螺旋状,失去了对称形,如螺类。
(二)外套膜(mantle)为身体背侧皮肤褶向下伸展而成,常包裹整个内脏团。外套膜与内脏团之间形成的腔称外套腔(mantle cavity)。腔内常有鳃、足,以及肛门、肾孔、生殖孔等开口于外套腔。
外套膜由内外两层上皮构成(图9-2),外层上皮的分泌物,能形成贝壳;内层上皮细胞具纤毛,纤毛摆动,造成水流,使水循环于外套腔内,借以完成呼吸、排泄、摄食等。左右2片套膜在后缘处常有一二处愈合,形成出水孔(exhalant siphon)和入水孔(inhalant siphon)。有的种类出入水孔延长成管状,伸出壳外,称为出水管和入水管。

(三)贝壳(Shell)体外具贝壳为软体动物的重要特征,因此研究软体动物的科学又称贝类学(Malacology)。大多数软体动物都具有一二个或多个贝壳,形态各不相同。有的呈帽状;螺类为螺旋形;掘足类为管状;瓣鳃类为瓣状。有些种类的贝壳退化成内壳,有的无壳。贝壳有保护柔软身体的功能。
贝壳的成分主要是碳酸钙和少量的壳基质(conchiolin或称贝壳素)构成。这些物质是由外套膜上皮细胞分泌形成的。贝壳的结构一般可分为3层(图9-2),最外一层为角质层(perios-tracum),很薄,透明,有光泽,由壳基质构成,不受酸碱的侵蚀,可保护贝壳。中间一层为壳层(ostracum),又称棱柱层(primatic layer),占贝壳的大部分,由角柱状的方解石(calcite)构成。最内一层为壳底(hypostracum),即珍珠质层(pearl layer),富光泽,由叶状霰石(aragonite)构成。外层和中层为外套膜边缘分泌形成,可随动物的生长逐渐加大,但不增厚;内层为整个套膜分泌而成,可随动物的生长而增加厚度。珍珠就是由珍珠质层形成的。当外套膜受到微小砂粒等异物侵入刺激,受刺激处的上皮细胞即以异物为核,陷入外套膜的上皮之间结缔组织中,陷入的上皮细胞自行分裂形成珍珠囊,囊即分泌珍珠质,层复一层地将核包住,逐渐形成珍珠。据史料记载,公元前2200多年,我国就有淡水育珠的记载(书经·禹贡篇),广西合浦育珠自古就很有名,采捕开始于汉代。
角质层和棱柱层的生长非连续不断的,由于食物、温度度等因素影响外套膜分泌机能,故贝壳的生长速度是不同的,因此在贝壳表面形成了生长线,表示出生长的快慢。
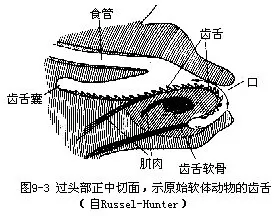
(四)消化系统 软体动物的消化管发达,少数寄生种类(内寄螺Entocolax)退化。多数种类口腔内具颚片(mandible)和齿舌(radula),颚片一个或成对,可辅助捕食。齿舌是软体动物特有的器官,位口腔底部的舌突起(odontophore)表面,由横列的角质齿组成,似锉刀状(图9-3)。摄食时以齿舌作前后伸缩运动,刮取食物。齿舌上小齿的形状和数目,在不同种类间各异,为鉴定种类的重要特征之一。小齿组成横排,许多排小齿构成齿舌。每一横排有中央齿一个,左右侧齿一或数对,边缘有缘齿一对或多对。齿舌上小齿的排列,以齿式表示,如中国圆田螺(Cipangopaludina chinensis)的齿式为 2·1·1·1·2。
(五)体腔和循环系统 软体动物的次生体腔极度退化,仅残留围心腔(pericardinal cavity)及生殖腺和排泄器官的内腔。初生体腔则存在于各组织器官的间隙,内有血液流动,形成血窦(blood sinus)。
循环系统由心脏、血管、血窦及血液组成。心脏一般位内脏团背侧围心腔内,由心耳和心室构成。心室一个,壁厚,能搏动,为血循环的动力;心耳一个或成对,常与鳃的数目一致。心耳与心室间有瓣膜,防止血液逆流。血管分化为动脉和静脉。血液自心室经动脉,进入身体各部分,后汇入血窦,由静脉回到心耳,故软体动物为开管式循环。一些快速游泳的种类,则为闭管式循环。血液无色,内含有变形虫状细胞。有些种类血浆中含有血红蛋白(haemoglobin)或血青蛋白(haemocyanin),故血液显红色或青色。
(六)呼吸器官 水生种类用鳃呼吸,鳃为外套腔内面的上皮伸展形成,位腔内。鳃的形态各异,鳃轴两侧均生有鳃丝,呈羽状,称楯鳃;仅鳃轴一侧生有鳃丝,呈梳状,称栉鳃(ctenidium);有的鳃成瓣状,称瓣鳃(lamellibranch);有些种类的鳃延长成丝状,称丝鳃(filibranch)。有的本鳃消失,又在背侧皮肤表面生出次生鳃(secondary branchium),也有的种类无鳃。鳃成对或为单个,数目不一,少则一个或一对,多则可达几十对。陆地生活的种类均无鳃,其外套腔内部一定区域的微细血管密集成网,形成肺,可直接摄取空气中的氧。这是对陆地生活的一种适应性。
(七)排泄器官 软体动物的排泄器官基本上是后肾管,其数目一般与鳃的数目一致,只有少数种类的幼体为原肾管。后肾管由腺质部分和管状部分组成,腺质部分富血管,肾口具纤毛,开口于围心腔;管状部分为薄壁的管子,内壁具纤毛,肾孔开口于外套腔。后肾管不仅可排除围心腔中的代谢产物,也可排除血液中的代谢产物。另外围心腔内壁上的围心腔腺,微血管密布,可排除代谢产物于围心腔内,由后肾管排出体外。
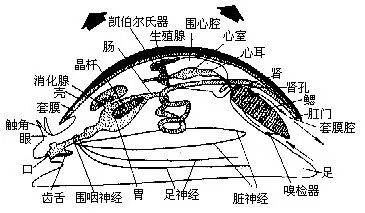
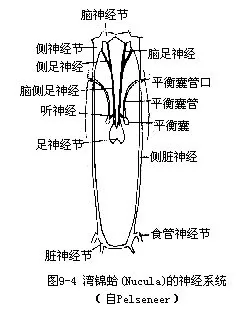
(八)神经系统 原始种类的神经系统无神经节的分化,仅有围咽神经环及向体后伸出的一对足神经索(pedal cord)和一对侧神经索(pleural cord)(图9-9)。较高等的种类,主要有4对神经节,各神经节间有神经相连。脑神经节(cerebral ganglion)位食管背侧,发出神经至头部及体前部,司感觉;足神经节(pedal gan-glion)位足的前部,伸出神经至足部,司运动和感觉;侧神经节(pleural ganglion)发出神经至外套膜及鳃等;脏神经节(visceralganglion)发出神经至各内脏器官(图9-4)。这些神经节有趋于集中之势,有的种类的主要神经节集中在一起形成脑,外有软骨包围,如头足类。软体动物已分化出触角、眼、嗅检器及平衡囊等感觉器官,感觉灵敏。
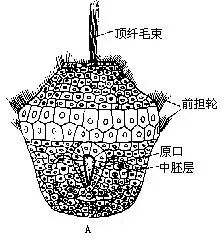
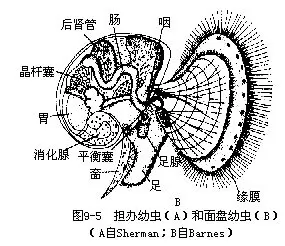
(九)生殖和发育 软体动物大多数为雌雄异体,不少种类雌雄异形;也有一些为雌雄同体。卵裂形式多为完全不均等卵裂,许多属螺旋型。少数为不完全卵裂。个体发育中经担轮幼虫和面盘幼虫(veliger larva)两期幼虫(图9-5),担轮幼虫的形态与环节动物多毛类的幼虫近似,面盘幼虫发育早期背侧有外套的原基,且分泌外壳,腹侧有足的原基,口前纤毛环发育成缘膜(velum)或称面盘。也有的种类为直接发育。淡水蚌类有特殊的钩介幼虫(glochidium)。
第二节 软体动物门的分类
软体动物目前已记载130000多种,分布广泛。依据它们的贝壳、足、鳃、神经及发生特点等特征,分为7纲。
一、单板纲(Monoplacophora)
绝大多数为化石种,已绝灭了近4亿年。这类动物有一个帽状或匙形的贝壳,有2~8对对称的肌痕,多年来一直被认为是腹足类中最原始的类(Patellacea)。1952年丹麦海洋研究船在太平洋沿岸哥斯达黎加(Costa Rica)3570m深海处第一次采得生活标本,1957年被定名为新碟贝(Neopilina galathea Lemche),以后又在大西洋、印度洋等处发现了一些种类。
新碟贝是一类原始的贝类,体为两侧对称,具一近圆形而扁的贝壳(图9-6B),腹足强大,周缘肌肉发达,中央薄,故无吸附能力,仅适于在海底滑行。缩足肌8对,分节排列于足的周围。足四周为外套沟,两侧共有鳃5或6对。足前端为口,后端为肛门。口前有一对具纤毛的口盖(ve-lum),口后有扇状触手一对(图9-6A)。心脏位围心腔内,由一心室及2对心耳构成;肾6或7对;雌雄异体,生殖腺2对,有生殖导管,开口于肾,生殖细胞由肾排出体外。神经系统由围食道神经环及向后伸出的侧神经和足神经组成。
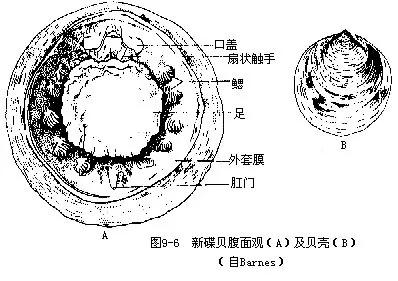
这类被称为“活化石”的原始贝类的发现,对研究贝类的起源与演化提供了新的资料。
二、无板纲(Aplacophora)
为软体动物中的原始种类,体呈蠕虫状,细长或短粗,无贝壳。体表被具石灰质细棘的角质外皮,头小,口在前端腹侧,躯体细长,腹侧中央有一腹沟,有的种类沟中有一小形具纤毛的足,有运动功能。体后有排泄腔,多数种类在腔内有一对鳃,腔后为肛门。无板类无触角、眼等感觉器官;肠为直管状,齿舌有或无;心脏为一心室一心耳,血管系统退化;雌雄同体或异体,个体发生中有担轮幼虫期。
无板类约有200多种,生活在低潮线下数10m至深海海底,肉食性,分布遍及全球。如毛皮贝(Chaetoderma)、新月贝(Neo-menia)、龙女簪(Proneomenia)(图9-7)等。我国南海海域79m深处曾采得龙女簪一种。

三、多板纲(Polyplacophora)
全部生活在沿海潮间带,常以足吸附于岩石或藻类上。体呈椭圆形,背稍隆,腹平。背侧具8块石灰质贝壳,多呈覆瓦状排列(图9-8A)。前面一块半月形,称头板(cephalic plate),中间6块结构一致,称中间板(intermediate plate),末块为元宝状,为尾板(tail plate),各板间可前后抽拉移动,因此动物脱离岩石后,可以曲捲起来。贝壳周围有一圈外套膜,称环带(girdle),其上丛生有小针、小棘等,形态各异。头部不发达,位腹侧前方,圆柱状,有一向下的短吻,吻中央为口。足宽大,吸附力强,在岩石表面可缓慢爬行。足四周与外套之间有一狭沟,即外套沟,在沟的两侧各有一列楯鳃,6对或数十对(图9-8B)。

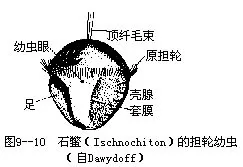
口腔具齿舌,消化腺发达。口腔前有一对唾液腺,食管后有一对粗大的糖腺,胃周围为肝。次生体腔发达,这也表现出其原始性。一心室,管状,二心耳。排泄为后肾管一对,肾口开于围心腔,肾孔开于外套沟中,后2对楯鳃之间。雌雄异体,具生殖腺和生殖导管,生殖孔开口于外套沟内(图9-8C)。受精卵经完全不均等卵裂,经囊胚,以内陷法形成原肠胚,后生出二纤毛带,发育成担轮幼虫(图9-10)、面盘幼虫,后体逐渐延长,腹面生足,背侧生壳,并具眼,经变态成成体。神经系统较为原始,由环食道的神经环与向后伸出的侧神经索和足神经索组成(图9-9)。神经索间有许多细神经相连,呈梯状。两侧神经索发出神经至外套,鳃及内脏器官;二足神经索主要发出神经至足部。有的种类贝壳上常有微眼(aesthetes),可感光。

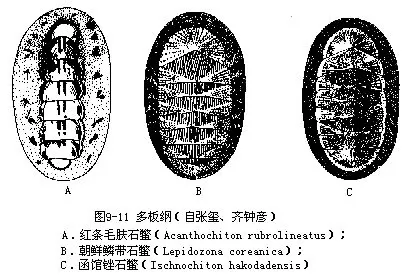
多板类约有1000种,我国沿海习见种类有:毛肤石鳖(Acanthochiton)体型较大,环带上有成丛的白色针束;鳞带石鳖(Lepidozona)壳片高,头板具放射肋;锉石鳖(Ischnochiton)具贝壳侧部有细肋(图9-11)。
四、腹足纲(Gastropoda)
软体动物中最大的一类,有10万种以上。生活在海洋、淡水及陆地,分布遍及全球,少数种类为寄生(内寄螺Entocolax、光螺Melanella、内壳螺Entoconcha等)。
(一)代表动物——圆田螺(Cipangopaludina) 圆田螺属为淡水中习见的大型螺类,分布较广,栖于湖泊、池沼、河流、水库、水田等处,以宽大的肉质足在水底爬行,以水生植物叶片、藻类等为食。中国圆田螺(C.chinensis)及中华圆田螺(C.cathayensis)等在我国分布甚广,前者为世界性种。
1.外部形态 圆田螺的壳大,薄而坚固,壳高50mm~60mm,宽35mm~40mm,呈圆锥形,螺层很膨胀,有6~7层。缝合线深,体螺层上生长线明显。
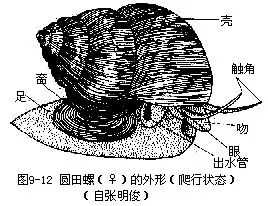
圆田螺的头和足可自壳口伸出,内脏团则留在壳内(图9-12)。头部发达,前端有一圆形突起称吻,吻腹侧为口。于吻基部两侧生有一对长圆锥形的触角,雄性右触角较左触角短而粗,有交接器的功能。触角的基部外侧各有一突起,其上各有一黑色眼。头后方两侧有褶状颈叶,右侧的发达,卷成管状,称出水管;左侧较小,贴在外套膜上,形成入水管。
头后身体腹面为宽阔的叶状足,肉质,前缘较平直,后端较狭。足背侧为内脏团,后部背面有一卵形的角质厣,其上有同心环形生长纹。当圆田螺缩入壳内时,首先头缩入,继而足跖面中央横折也缩入,厣正好封住壳口。
圆田螺的外套膜呈薄膜状,将内脏团包围,套膜边缘较厚,围绕头及足的周围,背缘及侧缘游离,腹缘与足愈合。在头足部与内脏团之间形成外套腔。
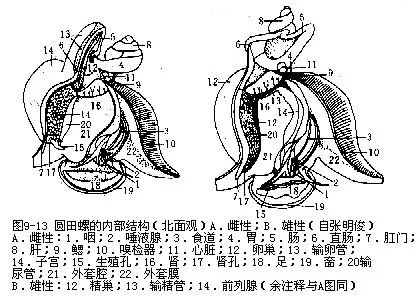
2.内部构造(图9-13)
(1)消化系统 口位吻前端腹面,内为膨大的咽,其内腔称口腔,具齿舌。咽后连以细的食管,伸至围心腔下,通入膨大的胃,咽与食管相连处的背侧,脑神经节之上,为二唾液腺,有导管自咽后两侧开口于口腔。唾液腺分泌粘液,无消化作用。胃周围为肝脏,为分支的管状腺组成,有肝管通入胃,能分泌醣酶和蛋白酶,是圆田螺的主要消化腺。胃后为肠,肠扭转180度,复向前伸,与输卵管平行,肛门开口于外套腔肾孔的右侧。
(2)呼吸器官 鳃栉状,一个,位外套腔左侧。鳃的上皮细胞具纤毛,内有血管。鳃正位于入水管内侧,当水流经过外套腔时,可摄取溶于水中的氧,排出二氧化碳。
(3)循环系统 由心脏和血管组成。心脏位于胃和肾之间的薄膜状围心腔内,由一心室和一心耳构成。心室壁厚,位后方;心耳壁薄,位前方,二者间有瓣膜。出鳃静脉连于心耳,心室伸出一主动脉,后分2支,一为头动脉,分布于头,外套膜和足等处。另一支为内脏动脉,分支到体后部内脏器官。各血管末端连于血窦。血液回心耳有二途径:一为经肾入鳃回到心耳,一为直接入鳃,回心耳,完成排泄和呼吸作用。圆田螺血液无色,含有变形虫状细胞。
(4)排泄系统 肾一个,略呈三角形,浅褐黄色,位围心脏之前,直肠左侧。肾右侧为一薄壁的输尿管,有孔与之相通。输尿管右侧壁与生殖器官(子宫或精巢)的外壁愈合。肾孔开口于肛门左侧稍后处,正位出水管的内侧,这样可使排泄物随水排出体外。
(5)神经系统 由神经节和神经连索构成,主要神经节4对(图9-14)。脑神经节一对,较大,二者有神经相连,位咽背侧,分出10对神经到触角、眼、口等体前部。侧神经节一对,位脑神经节之后,较小,左右不对称,由脑侧神经连索与脑神经节相连,以侧足神经连索连于足神经节。足神经节一对,长带状,位足的跖面中央处,两神经节有神经相连,每一足神经节分出神经至足前部和后部,由脑足神经连索连于脑神经节。脏神经节一对,形小,位食管末端处,彼此有神经相连。左脏神经节有一长神经连索与食管左侧的肠上神经节相连,肠上神经节又连于右侧神经节,其间的神经连索在食管上方,自左后至右前;右脏神经节也有一长神经连索与食管右侧的肠下神经节相连,肠下神经节又连于左侧神经节,其间神经连索在食管下方,自右后至左前。因此侧脏神经节间的神经连索于食管上下左右交叉形成“8”字形。
圆田螺感官发达,除皮肤有感觉作用外,触角为一感觉器官,其顶端有感觉细胞及神经末梢分布,感觉灵敏。平衡囊位足神经节内侧,为皮肤内陷的囊,上皮具纤毛,杂有感觉细胞,囊内有细小的耳石(otolith)。平衡囊分布有脑神经节分出的神经,可维持身体平衡。眼为视觉器官,也为皮肤内陷形成,具感觉细胞和色素细胞构成的视网膜,并有晶体。嗅检器为皮肤突起,是化学感受器,位于鳃近端部左侧,呈弯曲线状,色黄。
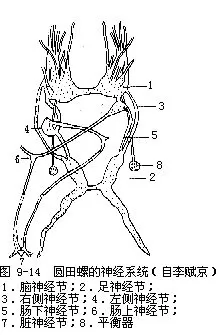
圆田螺的神经感官发达,与其活动的生活方式相适应。(6)生殖系统 田螺为雌雄异体,雄性右侧触角较雌性右侧触角粗大,此为鉴别雌雄的特征。雄性具精巢一个,较大,呈新月状,位外套腔右侧。精巢后端左侧连一输精管(在输尿管下方),向左横行,较短,后向前伸,膨大成贮精囊(前列腺),最后变细成射精管,入右侧触角中,其顶端的开口为雄性生殖孔。雄性的右触角有交接器的作用。雌性有卵巢一个,细长带状,黄色,与直肠上部平行。输卵管较短,连于卵巢,后端膨大通入子宫。子宫位于右侧,为一腺质壁的大形薄囊,可分泌蛋白质液包裹卵。子宫末端变细成管状,顶部为雌性生殖孔,位肾孔的右侧。田螺为体内受精,受精卵在子宫内发育生长,生下即为幼螺。雄田螺的右侧触角特化成交接器,卵胎生,这是腹足类中所特有的。
(二)主要特征 腹足类多营活动性生活,头部发达,具眼、触角。足发达,叶状,位腹侧,故称腹足类。足具足腺,为单细胞粘液腺。体外多被一个螺旋形贝壳,故又称单壳类(Univalvia)或螺类,有些种类为内壳或无壳。
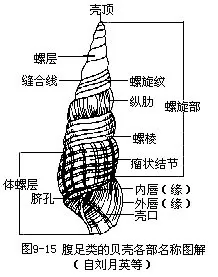
腹足类的贝壳形态为分类的重要依据。壳呈螺旋形,多数种类为右旋(dextral),少数左旋(senistral)。壳可分为2部分,含捲曲内脏器官的螺旋部(spire)和壳的最后一层,容纳头和足的体螺层(body whorl)。螺旋部一般由许多螺层(spiral whorl)构成,有的种类退化(鲍、宝贝等)。壳顶端称壳顶(apex),为动物最早形成的一层,各螺层间的界限为缝合线(suture),深浅不一。体螺层的开口称壳口(aperture),壳口内侧为内唇,外侧为外唇(图9-15)。壳口常有一盖,称厣(oper culum),角质或石灰质,为足的后端分泌形成,可封闭壳口。有些种类无厣(肺螺类)。螺轴为整个贝壳旋转的中轴,位贝壳内部中央,轴的基部遗留的小窝为脐(umbilicus),深浅不一。有的种类由于内唇外转而形成假脐(如红螺Rapana)。
口腔内常具齿舌和颚片;消化腺有唾液腺,是一种粘液腺,无消化作用;肝脏发达,为重要消化腺,可分泌醣酶及蛋白酶。有的种类肝脏尚有排泄功能(肺螺类)。
鳃一般呈栉状,一个,但原始种类为楯鳃[鲍、翁戎(Pleurotomaria)等];有些本鳃消失,生有次生鳃;陆生种类无鳃,以肺呼吸。
心脏具一心室,一或二心耳;肾一个,原始类型为一对。雌雄异体或雌雄同体。完全均等卵裂,属螺旋型,经有腔囊胚,以外包或内陷法形成原肠胚,有担轮幼虫和面盘幼虫。
神经系统由脑、足、侧、脏4对神经节组成,感觉器官有触角、眼、嗅检器(osphradium)、味蕾、平衡囊等,寄生种类无明显神经系统,感官极度退化或消失。
(三)身体不对称的起源 腹足类的头部和足表现出明显的两侧对称,内脏团呈螺旋形,失去对称形。这是因为在个体发生中身体经扭转的结果。
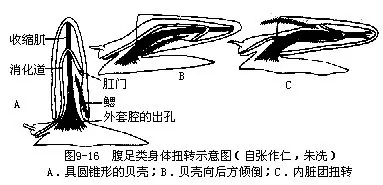
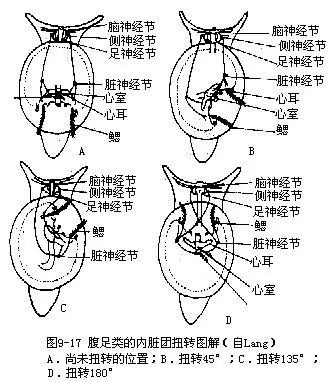
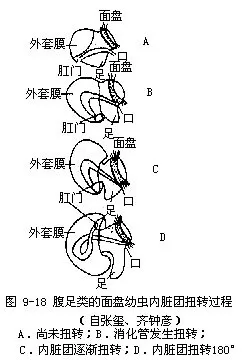
腹足类的身体不对称的起源问题很复杂,目前还没有很好解决。经古生物学的研究,下寒武纪(距今约5.5亿年)腹足类的化石,身体为左右对称。同时腹足类的个体发生中的担轮幼虫期,身体也是左右对称的,只是到了面盘幼虫的后期,才发生扭转,使身体逐渐失去对称形,这说明其祖先的身体是左右对称的。软体动物除腹足类外,其他类群的身体都是左右对称。这些都明显示出腹足类身体不对称是由左右对称的祖先经漫长的演变发生的,由于什么因素促使它们发生了这样的变化,目前一般解释是这样的:根据古生物学、胚胎学和比较解剖学的研究,认为腹足类的祖先身体为左右对称,心耳、鳃、肾等器官成对,左右对称排列,口在前端,肛门位体末。背侧有一腕形的贝壳,以腹面的足在水底爬行,与单板纲及多板纲相似。这种体制与已发现的化石标本是相符的。这样的腹足类动物,当遇到敌害时,则将身体缩入贝壳内,以保护自己。由于腹足逐渐发达,贝壳相应地也慢慢增大,不断向上方发展,成为圆锥形,与掘足类近似(图9-16A)。如此贝壳的容积增大了,口径小了,身体可以完全缩入壳内。在下寒武纪地层中发现的腹足类化石中,有不少种类就是这样的。但这样的贝壳有碍动物的运动,爬行中受到的阻力较大,也难以保持身体平衡。在演化发展中,高耸的贝壳逐渐向体后方倾倒(图9-16B),如此虽克服了爬行中的阻力,但使外套腔出口受压,肛门及肾孔等压在足和壳之间,影响水的循环,阻碍了各器官的正常生理机能。于是发生了适应的变化,身体的内脏团部分沿纵轴发生了扭转,内脏团扭转180°,肛门移到体前方,心耳、鳃、肾等器官左右易位。同时也发生螺旋曲捲,内脏团形成螺旋形。如此外套腔开口转移到体前,不再受压,水在腔内循环流畅,排泄作用等得以正常进行(图9-16C)。这样在贝壳的容积不变的情况下,其表面积减小了,从而爬行中受到水的阻力也减小。这一切变化对腹足类的生活是有利的,在长期演化发展中被保留下来。因为内脏团顺时针方向或逆时针方向的扭转,致使一侧的器官消失了,心耳、鳃、肾等均成为单个;侧神经节和脏神经节间的侧脏神经连接从平行而扭成“8”字形(图9-17)。这种扭转过程在腹足类个体发生中看得很清楚(图9-18)。腹足类这个扭转过程,自寒武纪开始至奥陶纪末期才完成,经历了千万年的演化过程。腹足纲中的后鳃类,其侧脏神经连接并不扭成“8”字形,身体表现为左右对称,这是它们又发生了逆扭转的结果。因已发生过扭转,故身体一侧器官已经消失。
(四)腹足纲分类 本纲约有10万以上,分3亚纲
1.前鳃亚纲(Prosobranchia)(扭神经亚纲Streptoneura)具外壳;头部具一对触角;鳃位心室前方;侧脏神经连索左右交叉成“8”字形。
原始腹足目(Archaeogastropoda):鳃楯状,一对;一般一心室二心耳;二肾;齿舌的角质齿一般极多。
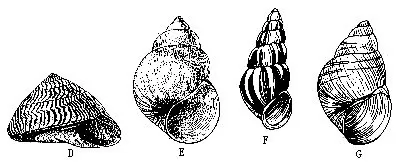
鲍(Haliotis)贝壳大而低,螺旋部退化;壳口极大;无厣;足肥大。为海味中的珍品,我国已进行人工养殖。壳可入药,称石决明。笠贝(Notoacmea)壳低圆锥形,如笠状,无螺旋部,无厣。蝾螺(Tur-bo)壳螺旋形,厣石灰质,厚。马蹄螺(Troc hus)壳圆锥形,珍珠质层厚,为制扭扣的优质原料。螺(Umbonium)壳圆锥形,较低而宽,壳质坚实,华北沿海潮间带沙滩习见种类(图9-19A~D)。
中腹足目(Mesogastropoda):栉状鳃一个;一心耳;一肾;通常无吻无水管种类多,有寄生种类。
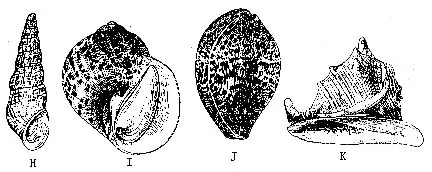
圆田螺。滨螺(Littorina)壳小,近球形,分布于海滨岩石上,退潮后可暴露在空气中生活。钉螺(Oncomelania)形似螺钉,为血吸虫的中间宿主,淡水产。沼螺(Parafossarulus)壳短圆锥形,厣石灰质,淡水产,中华枝睾吸虫的中间宿主。黑螺(Melanoides)壳长锥形,缝合线浅,卫氏并殖吸虫的中间宿主,淡水生活。玉螺(Natica)壳近球形,体螺层膨大,牡蛎养殖之敌害。内寄螺无壳,体退化呈筒状,寄生在棘皮动物体上。宝贝(Cypraea)壳的螺旋部退化,极小,埋于体螺层内。色鲜艳,富光泽,我国南海估计约有50种。冠螺(Cassis)大型螺。螺旋部小,体螺层膨大,整个贝壳呈冠状,壳质重厚。我国海南、台湾有分布,为螺类中最大种类。虎斑宝贝(C.tigris)和唐冠螺(C.cornuta)列为国家二级保护动物。水字螺(Pterocera chiragra)外唇极度扩张,生出6条匀称的长棘状突起,如“水”字状。海南、西沙群岛有之(图9-19E~L)。
新腹足目(Neogastropoda)(狭舌目Stenoglossa):栉鳃一个;一心耳;一肾;吻发达,有水管;齿舌狭长,小齿每排3个。全海产。
红螺壳陀螺形,大而厚;外唇内面呈红色,危害牡蛎养殖。荔枝螺(Thais)壳塔状,厚而坚,也为牡蛎养殖的敌害。骨螺(Murex)壳的各螺层上常具各种形状的突起,为贝类养殖之害。延管螺(Magilus)壳小,能分泌石灰质长管,栖于珊瑚礁间,我国海南有分布。织纹螺(Nassa)壳塔状,螺肋明显,生长线细。芋螺(Conus)壳似纺锤形,广东、海南有产。有的种类如织锦芋螺(C.textile)有毒腺,人被伤,严重时有生命危险(图9-19M~P)。



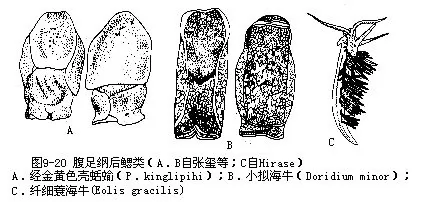
2.后鳃亚纲(Opisthobranchia)(直神经亚纲Euthyneura)贝壳不发达,有的为内壳(被鳃类),有的壳退化(无腔类),有的无壳(裸鳃类);触角一对、2对或无触角;鳃位心室后方;侧脏神经连索不左右交叉成“8”字形。主要的目有:
被鳃目(Tectibranchia):一鳃,多少为侧足或外套膜遮盖;一嗅检器;具外壳或内壳,无厣。
壳蛞蝓(Philine)体呈蛞蝓状,壳薄,具2螺层,被外套膜遮盖,完全包在内(图9-20A)。海兔(Aplysia)体肥满,形似兔,贝壳退化,小;2对触角。拟海牛(Doridium)触角一对,壳圆形,包在外套膜内,侧足发达(图9-20B)。
裸鳃目(Nudibranchia):无壳;无鳃;具2次性鳃。
蓑海牛(Eolis)蛞蝓状,体背侧有成列的锥状突起(图9-20C)。
3.肺螺亚纲(Pulmonata) 无鳃,以肺囊呼吸;多栖于陆地或淡水中;触角l~2对;贝壳无厣;直接发育。
基眼目(Basommatophore):具外壳;一对触角;眼位触角基部。
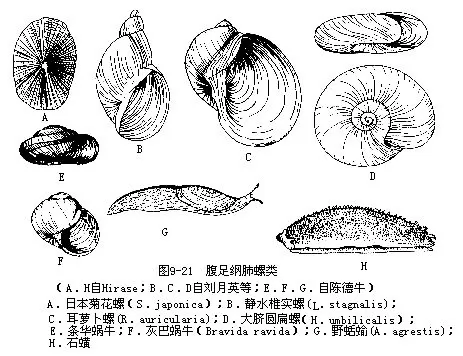
菊花螺(Siphonaria)贝壳锥形,有细的放射肋,生活于沿海潮间带。椎实螺(Lymnaea)壳薄,半透明,体螺层膨胀,无厣。萝卜螺(Radix)壳质薄,体螺层极膨胀,外形似耳状。此2种均为肝片吸虫的中间宿主。圆扁螺(Hippeutis)贝壳小,作水平旋转,扁平呈盘状,为姜片虫的中间宿主(图9-21A~D)。
柄眼目(Stylommatophore):贝壳发达或退化或无壳; 2对触角;眼位后触角的顶端。
华蜗牛(Cathaica)小型,壳呈低圆锥形。条华蜗牛(C.fasciola)壳黄褐色,体螺层有一条黄褐色带,全国分布。巴蜗牛(Bradybaena)贝壳扁球形,脐孔圆形,深,生活在潮湿山林间。蛞蝓(Agriolimax)体呈长叶状,具退化的内壳。世界性分布。石磺(Oncidium verruculatum)外形似海牛,无壳;肺囊退化,体背侧具枝状鳃,沿海有分布(图9-21E~H)。
五、掘足纲(Scaphopoda)
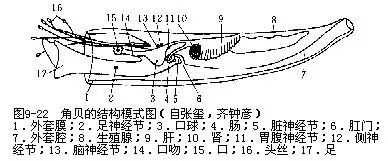
全海产。具长圆锥形稍弯曲的管状贝壳,如象牙状。粗的一端为前端,开口大,称为头足孔;细的一端为后端,开口小,称为肛门孔。壳凹的一面为背侧,凸的一面为腹侧。外套膜呈管状,前后端有开口。头部不明显,前端具有不能伸缩的吻,吻基部两侧生有许多头丝(captacula),能伸缩,末端膨大。头丝可伸出壳外,有触觉功能,也可摄食。掘足类为肉食性,吻内为口球,具颚片和齿舌。足在吻的基部之后,柱状,末端三叶状或盘状。足可伸得很长,能挖掘泥沙。肛门开口于足的基部腹侧(图9-22)。无鳃,以外套膜进行气体交换。循环器官心脏一室,无心耳,未分化出血管,仅有血窦。肾一对,囊状,位胃侧面。雌雄异体,生殖腺一个;个体发生中有担轮幼虫和面盘幼虫。
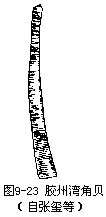
掘足类自潮间带至4000m深海都有分布,约300种,仅2科。角贝(Dentalium)壳呈象牙状,足圆锥形,末端有二翼状侧叶。本属动物在我国分布广,种类多。大角贝(D.vernedei)壳长100mm以上,厚;头足孔直径10mm左右,我国东海及南海由浅海至100m深处均有分布。胶州湾角贝(D.kiaochowwanensis)分布在青岛(图9-23)。
六、瓣鳃纲(Lamellibranchia)
瓣鳃纲动物全部生活在水中,大部分海产,少数在淡水,极少数为寄生(内寄蛤Entovalva、恋蛤Peregrinamor等)。约有2万种,分布很广。一般运动缓慢,有的潜居泥沙中,有的固着生活,也有的凿石或凿木而栖。
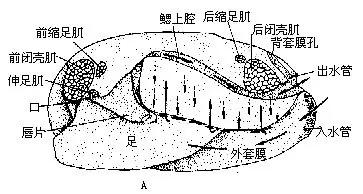
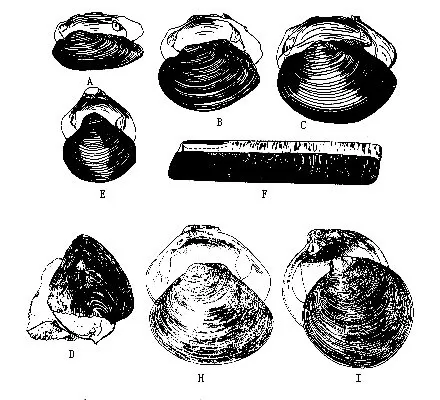
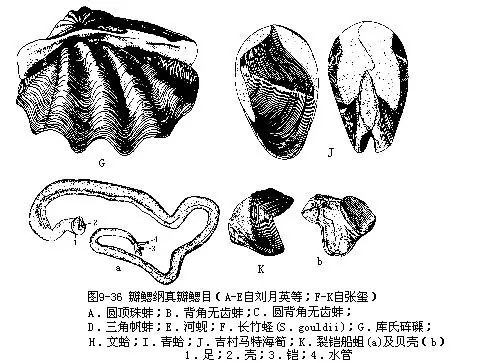
(一)代表动物——无齿蚌(Anodonta)无齿蚌又称河蚌,生活在淡水湖泊、池沼、河流等水底,半埋在泥沙中,体后端的出入水管外露,水可流入流出外套腔,借以完成摄食、呼吸及排出粪便、代谢产物等机能。河蚌滤食水中的微小生物及有机质颗粒等。背角无齿蚌(A.woodianawoodiana)及背圆无齿蚌(A.woodiana pacifica)在我国分布很广。
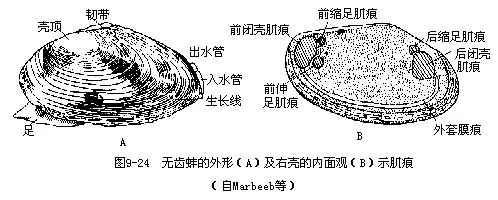
1.外部形态 无齿蚌具有两瓣卵圆形外壳,左右同形,壳顶突出。壳前端较圆,后端略呈截形,腹缘弧形,背缘平直。绞合部无齿,其外侧有韧带(图9—24A),依靠其弹性,可使二壳张开。壳面生长线明显。
壳的内面有肌肉附着的肌痕(图9-24B)。与壳腹缘并行的外套痕;壳前上方有3肌痕,最大的一个椭圆形,为前闭壳肌痕;其后上缘为一小的略呈三角形的前缩足肌痕;其后下缘为伸足肌痕。壳后端近背缘处有二肌痕,大的为后闭壳肌痕,椭圆形,其前上缘一小的是后缩足肌痕。
2.内部构造
(1)外套膜 紧贴二壳内面为两片薄的外套膜,包围蚌体,套膜间为外套腔。套膜内面上皮具纤毛,纤毛摆动有一定方向,引起水流。两片套膜于后端处稍突出,相合成出水管和入水管(图9-24A)。入水管在腹侧,口呈长形,边缘褶皱,上有许多乳突状感觉器;出水管位背侧,口小,边缘光滑。
(2)足呈斧状,左右侧扁,富肌肉,位内脏团腹侧,向前下方伸出。为蚌的运动器官。
(3)肌肉与壳内面肌痕相对应,可见前闭壳肌(anterioradductor)及后闭壳肌(posteriorad-ductor),为粗大的柱状肌,连接左右壳,其收缩可使壳关闭。前缩足肌(anterior retractor)、后缩足肌(posterior retractor)及伸足肌(protractor)(图9-25A),一端连于足,一端附着在壳内面,可使足缩入和伸出。
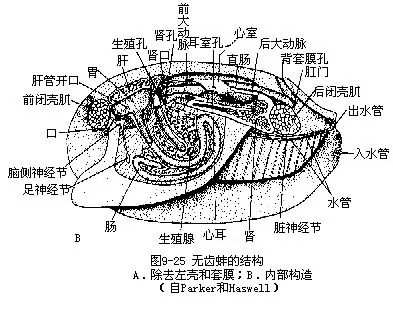
(4)消化系统 口位前闭壳肌下,为一横缝。口的两侧各有一对三角形唇片,大,密生纤毛,有感觉和摄食功能。口后为短而宽的食道,下连膨大的胃,胃周围有一对肝脏,可分泌淀粉酶、蔗糖酶,有导管入胃。胃后为肠,盘曲于内脏团中,后入围心腔,直肠穿过心室,肛门开口于后闭壳肌上,出水管附近(图9—25B)。胃肠之间有一晶杆,为一细长的棒状物,前端较粗,顶端形态变异较大,呈细尖、膨大、钩状、盘曲等。晶杆位于肠内,其前端突出于胃中,与胃楯下部相接。晶杆可能为储存的食物,河蚌在缺乏食物条件下,24小时后晶杆即消失,重新喂食,数天后晶杆恢复存在。河蚌以有机质颗粒,轮虫、鞭毛虫、藻类、小的甲壳类等为食。
(5)呼吸器官 在外套腔内蚌体两侧各具两片状的瓣鳃(lamina),外瓣鳃短于内瓣鳃。每个瓣鳃由内外二鳃小瓣(lamellae)构成,其前后缘及腹缘愈合成“U”形,背缘为鳃上腔(su-prabranchial chamber)。鳃小瓣由许多纵行排列的鳃丝(branchial filament)构成,表面有纤毛,各鳃丝间有横的丝间隔(interfilamental junction)相连,上有小孔,称鳃孔(ostrium)。二鳃小瓣间有瓣间隔(interlamellar junction),将鳃小瓣间的鳃腔分隔成许多小管,称为水管(water tube)。丝间隔与瓣间隔内均有血管分布,鳃丝内也有血管及起支持作用的几个质棍(chitinous rod)(图9-26C)。
由于鳃及外套膜上纤毛摆动,引起水流,水由入水管进入外套腔,经鳃孔到鳃腔内,沿水管上行达鳃上腔,向后流动,经出水管排出体外。水经过鳃时,即进行气体交换。外套膜也有辅助呼吸的功能。每24小时经蚌体内的水可达40L。鳃表面的纤毛可滤食水中的微小食物颗粒,送至唇片,再入口。因此鳃尚可辅助摄食。外瓣鳃的鳃腔又是受精卵发育的地方,直至钩介幼虫形成。

(6)循环系统 由心脏、血管、血窦组成。心脏位脏团背侧椭圆形围心腔内,由一长圆形心室及左右两薄膜三角形心耳构成(图9—25B,图9—27)。心室向前向后各伸出一条大动脉。向前伸的前大动脉(aorta)沿肠的背侧前行,后大动脉沿直肠腹侧伸向后方,以后各分支成小动脉(artery)至套膜及身体各部,最后汇集于血窦(外套窦、足窦、中央窦等),入静脉,经肾静脉入肾,排除代谢产物,再经入鳃静脉入鳃,进行氧碳交换,经出鳃静脉回到心耳。部分血液由套膜静脉入心耳,即外套循环(图9—27)。

无齿蚌血液中含血青蛋白(haemocyanin),氧化时呈蓝色,还原时无色,其与氧结合能力不及血红蛋白。一般软体动物100mL血液中含氧通常不超过3mg。血液中含变形虫状细胞,有吞噬作用。因此血液除输送养分外,尚有排泄功能。变形虫状细胞聚集,其伪足部分互相结合,使血液凝固(蚌血液中无纤维蛋白原)。
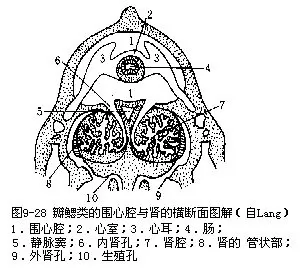
(7)排泄器官 蚌具一对肾,由后肾管特化形成,又称鲍雅诺氏器(organ of Bojanus);还有围心腔腺,亦称凯伯尔氏器(Keber's organ)。肾位于围心腔腹面左右两侧,各由一海绵状腺体及一具纤毛的薄壁管状体构成,呈“U”形。前者在下,肾口开于围心腔;后者在上,肾孔开口于内瓣鳃的鳃上腔前端(图9-25B,图9-28)。围心腔腺位围心腔的前壁,为一团分支的腺体,由扁平上皮细胞及结缔组织组成,其中富血液,可收集代谢产物,排入围心腔,经肾排出体外。各组织间的吞噬细胞,也有排泄功能。

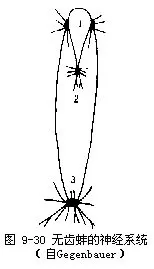
(8)神经系统 无齿蚌具有3对神经节(图9-29,图9-30)。前闭壳肌下方,食道两侧为一对脑神经节,很小,实为脑神经节和侧神经节合并形成,可称为脑侧神经节。在足的前缘靠上部埋在足内的为一对长形的足神经节,二者结合在一起(图9-25B)。脏神经节一对,已愈合,呈蝶状,位后闭壳肌的腹侧的上皮下面,较大(图9-30)。脑、足、脏3对神经节之间有神经连索相连接,脑脏神经连索较长,明显。
蚌的感官不发达,位足神经节附近有一平衡囊,为足部上皮下陷形成,内有耳石,司身体的平衡。脏神经节上面的上皮成为感觉上皮,相当于腹足类的嗅检器,为化学感受器。另外在外套膜、唇片及水管周围有感觉细胞的分布。
(9)生殖和个体发育 蚌为雌雄异体,生殖腺位足部背侧肠的周围,呈葡萄状腺体,精巢乳白色,卵巢淡黄色。生殖导管短,生殖孔开口于肾孔的后下方,很小(图9—25B,图9—28)。
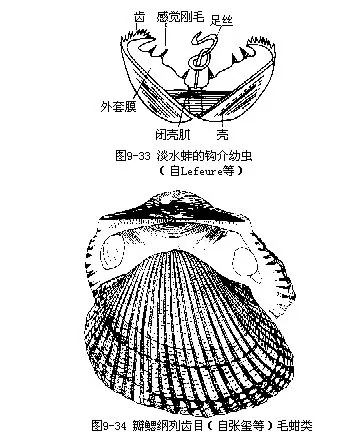
蚌的生殖季节一般在夏季,精卵在外瓣鳃的鳃腔内受精,受精卵由于母体的粘液作用,不会被水流冲出,而留在鳃腔中发育。故外瓣鳃的鳃腔又称育儿囊(marsupium)。经完全不均等卵裂(属螺旋型),发育成囊胚,以外包和内陷法形成原肠胚,发育成幼体,在鳃腔中越冬。观察背角无齿蚌的早期胚胎细胞,确定其染色体2n=38或2n=37(第19号染色体为单个)二者的差异不明,可能与性染色体有关,(马庆福等,1987)。来年春季,幼体孵出,发育成河蚌特有的钩介幼虫(相当于其他瓣鳃类的面盘幼虫)。幼虫具双壳,有发达的闭壳肌,壳的腹缘各生有一强大的钩,且具齿。腹部中央生有一条有粘性的细丝,称足丝(图9—33)。壳侧缘生刚毛,有感觉作用。幼虫有口无肛门。幼虫可借双壳的开闭而游泳。淡水中鳑鱼(Rhodaus sinensis)等,以长的产卵管插入蚌的入水管,产卵于蚌的外套腔中。如此蚌的钩介幼虫有机会接触鳑鱼,可寄生在鱼的鳃、鳍等处。鱼皮肤受其刺激而异常增殖,将幼虫包在其中,形成囊状。幼虫以外套膜上皮吸取鱼的养分。经2~5周,变态成幼蚌,破囊离鱼体,沉入水底生活。经5年方达性成熟。以后仍继续生长。
(二)主要特征 体具两片套膜及两片贝壳,故称双壳类(Bivalvia);头部消失,称无头类(Acephala);足呈斧状,称斧足类(Pelecypoda);瓣状鳃,故称瓣鳃类。
贝壳一对,一般左右对称,也有不对称的(不等蛤Anomia及牡蛎Ostrea等)。壳的形态为分类的重要依据。贝壳中央特别突出的一部分,略向前方倾斜,称为壳顶(umbo),这是壳中最老的部分。壳顶所在处,为壳的前方。相反的一端为后方。以壳顶为中心,有同心环状排列的生长线,有的种类有自壳顶向腹缘有放射的肋或沟。壳顶前方常有一小凹陷,称小月面,壳顶后的为楯面。壳的背缘较厚,于此处常有齿和齿槽,左右壳的齿及齿槽相互吻合,构成绞合部(hinge)。绞合齿的数目和排列不一,为鉴定双壳类种类的主要特征。绞合齿中正对壳顶的为主齿,其前的齿称前侧齿,其后为后侧齿。在绞合部连结两壳的背缘有一角质的、具弹性的韧带(ligament),其作用可使二壳张开。壳自背至腹为其高度,自前至后为其长度,两壳左右最宽处为其宽度(图9-31)。
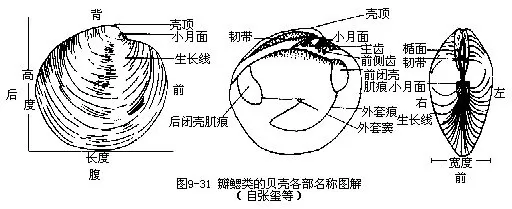
一些种类(贻贝、蚶、扇贝等)在足的腹中线稍后处有一孔,称为足丝孔,通入足丝囊内,其上皮细胞的分泌物遇水即变硬成贝壳素的丝状物,集合成足丝(byssus),用以固着外物。
口为上下二唇间的横缝,唇多为三角形,具纤毛,可摄食。胃肠间有晶杆(crystalline style),细长棒状。胃中有胃楯(gastric shield),有保护胃的作用。
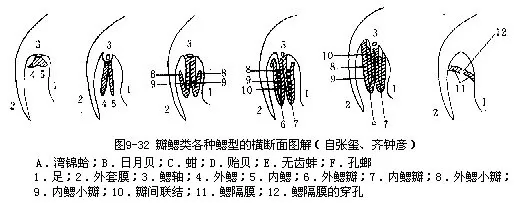
鳃在原始种类(湾锦蛤Nucula)为楯状;有的为丝状或瓣状;有的鳃瓣互相愈合,且退化,形成一有孔的隔膜,为隔鳃(孔螂类Poromyacea)(图9-32),已无呼吸作用。
心脏为一心室二心耳构成,开管式循环;排泄器官为一对肾;神经节有脑、足、脏3对,感官不发达。多数雌雄异体,少数雌雄同体(牡蛎),个体发生中有担轮幼虫及面盘幼虫。淡水蚌有特有的钩介幼虫(图9-26)。
(三)瓣鳃纲分类 瓣鳃纲约有2万种,依绞合齿的形态、闭壳肌发育程度和鳃的结构等,分为3目。
1.列齿目Taxodonta绞合齿多,同形,排成一列;闭壳肌2个,均发达。楯鳃或丝鳃。
湾锦蛤(Nuculo)壳小而厚,卵圆形;楯鳃小,鳃丝完全横列(图9-32A),我国黄渤海有分布。云母蛤(Yoldia)壳前方常开口,鳃丝直,不反折,黄渤海数十米深海底泥沙中有之。蚶(Ar-ca)壳厚,膨胀,壳面有粗的放射肋,鳃丝常反折(图9-32C)。如毛蚶(A.subcrenata)(图9-34)、泥蚶(A.granosa)、魁蚶(A.inflata)等为习见食用贝类,我国沿海均有分布。
2.异柱目Anisomyaria前闭壳肌很小或消失,后闭壳肌发达;绞合齿一般退化或成小结节状,或无绞合齿。鳃丝间以纤毛盘或结缔组织相连接。
贻贝(Mytilus edulis)壳略呈三角形,壳顶尖,腹缘平直。具足丝。其肉体干制品称淡菜,味鲜美。贻贝已大量进行人工养殖。栉孔扇贝(Chlamys farreri)壳呈扇状,放射肋明显,壳的前耳大于后耳。其后闭壳肌干制品称干贝,为海味中的上品。珍珠贝(Pteria)左右壳大小不等,无绞合齿,为生产珍珠的母贝。马氏珍珠贝(P.martensii)为世界著名的生产珍珠的母贝,我国广东、海南有分布。江瑶(Pinna)为大型种类,两壳等大,壳质脆,三角形。其闭壳肌的干制品称江瑶柱,为海味中的珍品。广东、海南、西沙群岛等地有分布。牡蛎(Ostrea)左壳大,略凹,固着外物;右壳小,平。无绞合齿;壳面有放射肋和鳞片层。为海产贝类中主要养殖种类,我国沿海有20多种。密鳞牡蛎(O.denselamellosa)壳近圆形,鳞片层较密,全国沿海均有分布。僧帽牡蛎(O.cucullata)壳小,三角形,为东南沿海重要养殖种类(图9-35)。
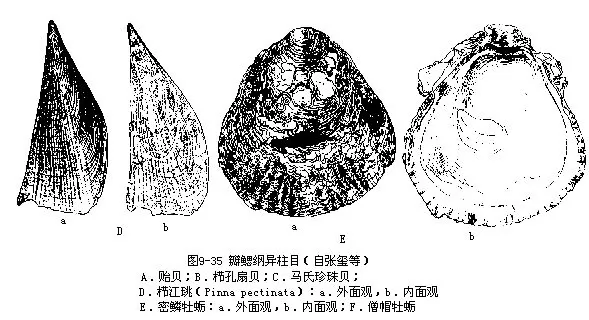
3.真瓣鳃目Eulamellibranchia铰合齿少或无;前后闭壳肌均发达,大小相等;鳃丝和鳃小瓣间以血管相连接;出水孔和入水孔常形成水管。
无齿蚌为淡水产,壳卵圆形,无绞合齿。我国有50多种。珠蚌(Unio)壳薄、长圆形。圆顶珠蚌(U.douglasiae)左右壳有拟主齿二,左壳侧齿二,右壳一,我国分布广、壳可入药。帆蚌(Hyriopsis)壳背缘向上伸出一很大的三角形帆状翼。三角帆蚌(H.cumingii)为淡水育珠的优良品种,已进行人工养殖,分布于华北各地。蚬(Corbicula)壳呈球形,绞合部主齿发达。河蚬(C.fluminea)壳近三角形,全国分布,肉可食用(图9-36A~E)。竹蛏(Solen)壳长形,薄,双壳合抱似竹筒状,足发达,为食用的佳品。砗磲(Tridacna)贝壳极大,重厚,库氏砗磲(T.coo-kiana)壳巨大,长超过1m,重200kg以上,为双壳类中最大的种类,列为国家一级保护动物。我国海南及西沙有分布。文蛤(Mertrix mertrix)壳近三角形,壳质厚,表面光滑,具花纹,肉可食,壳可作面油容器。青蛤(Cyclina sinensis)壳薄,近圆形,生活于浅泥滩,为食用蛤,肉鲜美(图9-36A~I)。海筍(Martesia)壳质薄,背缘反折在壳顶上;绞合部无齿。凿石而居,危害海港岩石建筑,我国沿海分布的吉村马特海蛆(M.yoshimurai)繁殖力强,危害甚大(图9-36J)。有的海筍(M.striata)穴居木材中。船蛆(Teredo)体呈蠕虫状,壳小而薄,球形,仅包住身体的前端一小部分,钻木而栖。船蛆繁殖力强,生长迅速,对沿海码头、木桩、木船等木建筑破坏严重。裂铠船蛆(T.manni)在我国沿海各地均有发现(图9-36K)。
七、头足纲(Cephalopoda)
头足类全海产,肉食性。体左右对称,分头、足、躯干三部分。头部发达,两侧有一对发达的眼;原始种类具外壳,多数为内壳或无壳;足着生于头部,特化成腕和漏斗,故称头足类(图9-37)。漏斗位于头腹面、头与躯干之间。羽状鳃一对或2对,心耳和肾的数目与鳃一致。具软骨。口腔有颚片和齿舌。神经系统集中,感官发达。闭管式循环系统。直接发生。
(一)代表动物——乌贼(Sepia)乌贼俗称墨鱼,渔业捕捞量很大,肉鲜美,富营养。生活在温暖海洋中,游泳快速,主要以甲壳类为食,也捕食鱼类及其他软体动物等。
1.外形身体可区分为头、足和躯干3部分,躯干相当于内脏团,外被肌肉性套膜(图9-38A),具石灰质内壳(图9-40)。
(1)头位体前端,呈球形,其顶端为口,口周围具口膜,外围有5对腕。头两侧具一对发达的眼,构造复杂。眼后下方有一椭圆形的小窝,称嗅觉陷,为嗅觉器官,相当腹足类的嗅检器,为化学感受器(图9-37)。
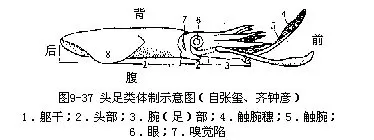
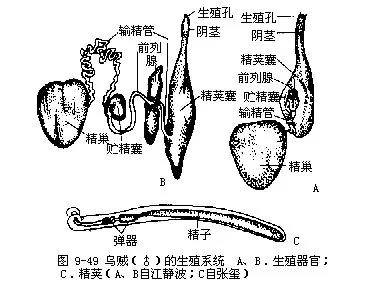
(2)足已特化成腕和漏斗。腕10条,左右对称排列,背部正中央为第一对,向腹侧依次为2~5对,其中第4对腕特别长,末端膨大呈舌状,称为触腕(tentacular arm),可以捕食,能缩入触腕囊内。各腕的内侧均具4行带柄的吸盘,触腕只在末端舌状部内侧有10行小吸盘,此称触腕穗(tentacular club)。雄性左侧第4腕的中间吸盘退化,特化为生殖腕或称茎化腕(hectocotylizedarm)。可输送精荚入雌体内,起到交配器的作用。根据茎化腕可鉴别雌雄。
漏斗位头的腹侧,基部宽大,隐于外套腔内,其腹面两侧各有一椭圆形的软骨凹陷称闭锁槽(adhering groove)。与外套膜腹侧左右的闭锁突(adhering ridge)相吻合,如子母扣状,称闭锁器(adhering apparatus),可控制外套膜孔的开闭(图9-38B)。漏斗前端呈筒状水管,露在外套膜外,水管内有一舌瓣,可防止水逆流。
当闭锁器开启,肌肉性套膜扩张,海水自套膜孔流入外套腔;闭锁器扣紧,关闭套膜孔,套膜收缩,压水自漏斗的水管喷出。此为乌贼运动的动力。
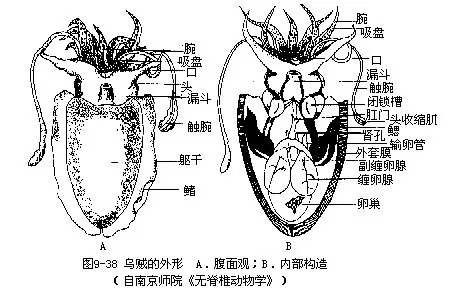
(3)躯干呈袋状,背腹略扁,位头后。外被肌肉非常发达的套膜,其内即为内脏团。躯干两侧具鳍,鳍在躯干末端分离。鳍在游泳中起平衡作用。由于躯干背侧上皮下具有色素细胞,可使皮肤改变颜色的深浅。
乌贼躯体方位依其在水中的生活状态,头端为前,躯干末端为后,有漏斗的一侧为腹,相反一侧为背。但根据软体动物的体制与乌贼的形态比较,其前端应为腹侧,因足位腹侧,后端为背,背侧为前,腹侧为后,这是乌贼的形态学位置。但为了观察叙述简便,多采用前种定位。
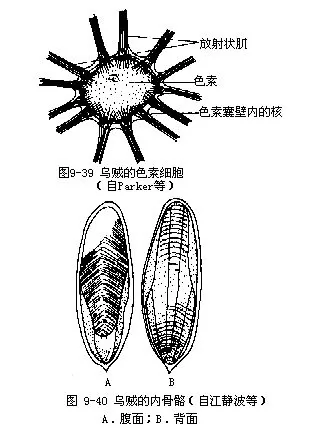
2.构造(1)体壁由上皮、结缔组织、肌肉组成,具内骨骼。上皮为单层细胞,其下有许多色素细胞(chromatophore),呈扁平状,细胞膜富弹性,周围有放射状的肌纤维(图9-39)。由于肌纤维的收缩,使色素细胞扩大呈星状,肌纤维舒张,色素细胞恢复原状,如此可使皮肤改变颜色的深浅。上皮下尚有一种虹彩细胞(iridocyste),使体表具光泽。
(2)内骨骼由内壳及软骨组成。内壳位于体背侧皮肤下的壳囊内,很发达,呈长椭圆形,前端圆,末端有一尖形突起(图9-40)。壳为石灰质,背侧硬,腹侧疏松,空隙多。内壳不但可以增加身体的坚强性,又可使身体比重减小,有利于游泳,并有助于保持平衡。软骨发达,其结构与脊椎动物相似,只是细胞有较长的分枝。主要软骨有头软骨,包围中枢神经系统和平衡囊,上具孔,神经可伸出。还有颈软骨、腕软骨等。
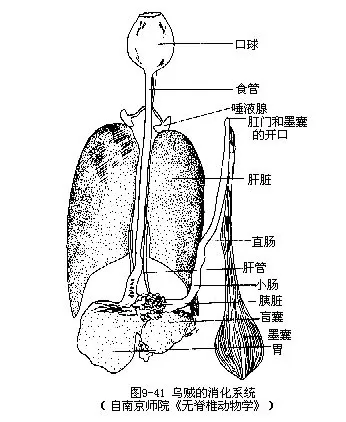
(3)消化系统乌贼的消化管呈“U”形。口位前端,口膜中央,口内为肌肉性口腔,称口球,其内有一对似鹦鹉喙状的颚片,一位背侧,一位腹侧,可切碎食物。口球底部为齿舌,可帮助吞咽食物。口腔内有前后唾液腺。前唾液为单个,唾液管开口于齿舌两侧,可分泌粘液;后唾液腺一对,位食管前端背侧,有导管通入口球,分泌毒液,可杀伤、麻痹捕获的动物。口球下接细长的食管,连于胃的贲门部。胃位内脏囊顶端,为长囊状,壁富肌肉。胃左侧为一盲囊,内壁褶皱,具纤毛。肠短而粗,自胃幽门部转向前伸,稍作拱曲,末端为直肠(图9-41),以肛门开口于外套腔,漏斗基部后方(图9-38B)。肛门两侧有一对肛门瓣,功能不详。
肝脏一对,甚大,为黄色腺体,占据内脏囊的前半部,位食管两侧。前端圆,后端尖。一对肝脏导管沿肠的两侧向后行,后两管会合,通入胃的盲囊。在肝脏导管上被有分支的腺体为胰脏(图9-41)。肝可分泌酶,输入胃中,进行消化作用。肝管有节律收缩,可自盲囊和胃中吸收养分,故有储存营养物质的功能。胰分泌淀粉酶及蛋白酶亦入胃中。消化后的食物入盲囊吸收,残渣由肛门排出体外。在直肠的末端近肛门处有一导管,连一梨形小囊,即墨囊(ink sac),位内脏团后端,实为一极发达的直肠盲囊(图9-42)。囊内腺体可分泌墨汁,经导管由肛门排出,使周围海水成墨色,借以隐藏避敌,乌贼之名来源于此。
(4)呼吸器官羽状鳃一对,位外套腔前端两侧(图9-38B)。每鳃有一鳃轴,两侧生有鳃叶,鳃叶是由许多鳃丝组成。鳃上密布微血管,水流经鳃,完成气体交换。鳃轴背缘有鳃腺,富血管,可能与鳃的营养有关。
(5)循环系统乌贼的循环系统基本为闭管式,仍有一些血窦。心脏由一心室二心耳组成,位体近后端腹侧中央围心腔内。心室菱形,不对称,壁厚,心耳长囊状,壁薄(图9-43)。心室向前伸出一前大动脉,分枝至头、套膜、消化管等处;心室向后伸出一后大动脉,至套膜、肾、直肠、生殖腺等器官。血液经微血管网汇入主大静脉,主大静脉分2支成肾静脉入肾;肾静脉及体后的外套静脉入鳃基部的鳃心(branchial heart)(鳃心壁为海绵质,可收缩),由入鳃静脉入鳃,再由出鳃静脉入左、右心耳,返回心室(图9-43)。血液循环中,在肾内排出代谢产物,在鳃内进行氧碳交换。头足类的血压甚高,可以超过脊椎动物。

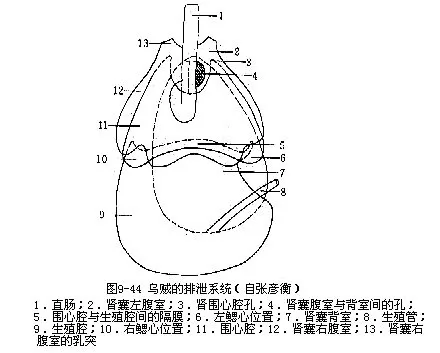
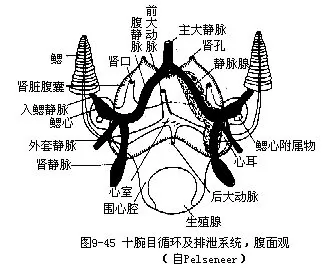
(6)排泄系统肾一对,为囊状结构,包括一背室和二腹室。二腹室位直肠背面两侧,左右对称(图9-44)。一对肾孔,开口于直肠末端两侧套膜腔中(图9-38B)。围心腔以一对导管伸入腹室,其开口为肾口。肾可自围心腔内收集代谢产物。二肾静脉周围有海绵状的静脉腺(图9-45),其分支中空,与静脉相通。这些腺体具有一层有排泄功能的腺质上皮,可从血液中吸收代谢产物,排入肾囊。肾的背室位于腹室的背侧,有孔与腹室相通。乌贼的排泄物不含尿酸,而是鸟嘌吟(guanin,C5H5ON5)。
(7)神经系统乌贼的神经系统发达,由中枢神经系统、周围神经系统及交感神经系统组成,结构复杂(图9-46)。
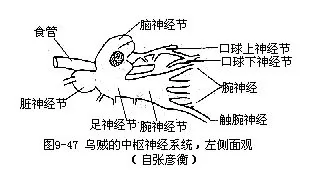
中枢神经系统由食管周围的脑神经节、脏神经节和足神经节等3对神经节组成(图9-47),外有一软骨质壳包围。食管背侧为一对脑神经节,腹侧为一对足神经节和一对脏神经节,二者前后排列。另有一对腕神经节,位足神经节前方,并与之相连。一对口球神经节位脑神经节前(图9-47)。
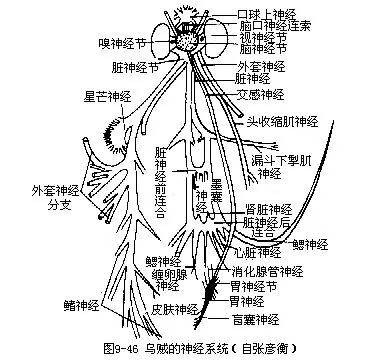
周围神经系统由中枢神经伸出的神经组成。脑神经节发出视神经,又分出嗅神经等;脏神经节伸出外套神经,其外枝于漏斗基部两侧形成一对星芒神经节,内枝分出皮肤神经及鳍神经等。又分出漏斗神经、头缩肌神经等(图9-46)。
交感神经由口球下神经节后面中央处分出的2条,沿食管两侧后行达于胃,形成胃神经节,卵圆形,位胃前端腹面。由此发出盲囊神经、胃神经、肠神经等(图9-46)。
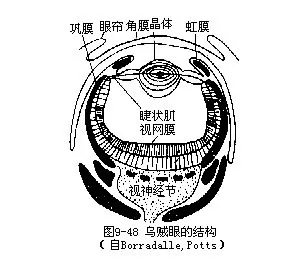
感官发达,有眼、平衡囊、嗅觉陷等。眼结构复杂(图9-48),最外为透明的角膜,无孔;中层为巩膜,瞳孔周围为虹彩,连于巩膜,瞳孔后为晶体和捷状肌;内层为视网膜,主要由杆状体组成,外层是视网膜细胞。眼的构造似脊椎动物,但由外胚层内陷形成。平衡囊一对,位头软骨内,介于足神经节和脏神经节之间。囊内充满液体,有一耳石,囊内前端背面有听斑(macula statica),另有突起称听脊(crista statica),为感觉作用部分。嗅觉陷位眼后下方,为上皮下陷,具有感觉细胞,脑神经节分出神经至此,为化学感受器。
(8)生殖系统乌贼为雌雄异体,外形上区别不明显。生殖为体外受精,直接发育。
雌性具卵巢一个,由体腔上皮发育形成,位内脏团后端生殖腔中(图9-38B)。卵成熟后落在腔内,由粗大的输卵管输出,管末端细,雌性生殖孔开口于鳃基部前方外套腔内。输卵管近末端处有一输卵管腺,其分泌物形成卵的外壳。直肠两侧内脏囊壁上为一对大的卵形缠卵腺(ni-damental gland),开口于外套腔,其分泌物也形成卵的外壳及一种遇水即变硬的弹性物质,可将卵粘成卵群。缠卵腺前还有一对小形副缠卵腺(图9-38B),功能不明。生殖季节时,卵分批成熟,分批产出。
雄性有精巢一个,位体后端生殖腔中,来源于体腔上皮。由许多小管集成,精子成熟后,由小管落入生殖腔中。输精管长,曲折一团,管上有精囊和摄护腺,端部膨大成精荚囊(spermatophoresac),末端为阴茎,雄性生殖孔开口于外套腔。精荚囊内有极多的精荚。精子到达精荚囊内,包被一层弹性鞘而形成精荚(图9-49)。
每年春夏之际,乌贼由深水游向浅水内湾处产卵,此谓生殖洄游。据青岛附近情况,产卵时适宜温度为15℃~20℃,盐分为30‰以上。产卵前雌雄交配,即雄性以茎化腕将精荚送入雌体外套腔中,精荚破裂,释放出里面的精子,精卵在外套腔内受精。交配后不久,雌性即排出受精卵,圆形,一端稍尖,长径10mm许,成串聚积一起,表面黑色,粘于外物上,俗称“海葡萄”。乌贼卵含大量卵黄,属端黄卵。经不完全卵裂(盘式卵裂),以外包法形成原肠胚,直接发育。孵化出幼体与成体相似。
我国乌贼种类较多,洄游明显的为曼氏无针乌贼(Sepiella maindroni),盛产于浙江南部沿海及福建沿海;台湾枪乌贼(Loligo formosana),分布于台湾海峡以南海区,汕头外海及北部湾为产卵场所。
(二)头足纲分类头足类的生活种类约有700种,化石种类在10000种以上。主要根据鳃和腕的数目分为2亚纲4目。
1.四鳃亚纲(Tetrabranchia)具外壳;腕数十个(60~90),无吸盘;漏斗为左右二叶组成; 2对鳃;2对心耳;2对肾。
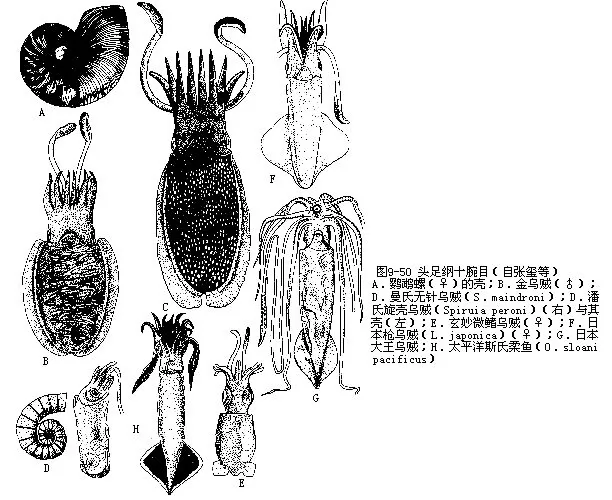
鹦鹉螺目(Nautiloidea):外壳的隔膜与壳壁结合的缝合线为直线,不曲折。约有3500种,绝大多数为化石种,生存种类仅存鹦鹉螺属(Nautilus)(图9-50A),共3种。鹦鹉螺生活在南太平洋热带海区,在数百m的海底营底栖生活,也可短暂的浮动和游泳。生殖期间由深海向浅海移动。鹦鹉螺(N.pompilius)为模式种,我国在海南、西沙和南沙均未采到生活标本,只获得空壳,长径达200mm。菲律宾群岛海域年采捕量达2万个。鹦鹉螺列为我国一级保护动物。
菊石目(Ammonoidea):缝合线曲折,极为复杂,全为化石种,约5000多种,可供划分地层及探矿时的参考。如菊石(Ammononites)、箭石(Belemnite)等。
2.二鳃亚纲 Dibranchia具内壳或无壳;腕8~10个,具吸盘;漏斗为完整的管子;二鳃;二心耳;二肾。
十腕目(Decapoda):5对腕,吸盘有柄;有石灰质内壳。金乌贼(Sepia esculenta)体卵圆,大 型,长可达200mm;内壳末端有骨针,为我国沿海习见种类。无针乌贼(Sepiella)内壳末端无骨针,体卵圆,长可达150mm,是我国产量最大的一种头足类,构成我国四大渔业之一,盛产浙江沿海。旋壳乌贼(S.spirula)具螺旋形石灰质内壳,裸露,暖海产。我国南海产旋壳乌贼。微鳍乌贼(Idiosepius)体小,鳍微小,位体末端,贝壳极退化。我国沿海产玄妙微鳍乌贼(I.paradoxa),体长20mm许。枪贼(Loligo)鳍三角形,占体长1/2以上,内壳角质。台湾枪乌贼(L.for-mosana)我国沿海产量很大,为食用种类。日本大王乌贼(Architeuthis japonica)体巨大,长可达1m以上,腕长4m,为无脊椎动物中最大者。柔鱼(Ommatostrephes)体略呈圆筒状,鳍三角形,不及体长1/2;内壳角质(图9-50B~H)。营养价值高,为食用头足类的上品。
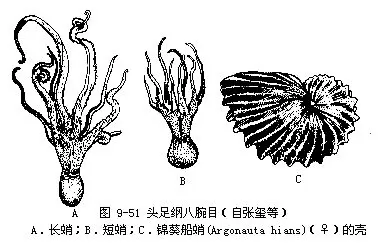
八腕目(Octopoda):体略呈球形;4对腕,吸盘无柄,腕间膜发达;内壳退化或完全消失。有须类(Cirrata)的外套膜背侧具一或2对鳍,腕上有须毛,深海产。无须类(Incirrata)无鳍,无须毛。章鱼(Octopus)体椭圆形,无鳍;右侧第3腕为茎化腕。长蛸(O.variabilis)腕长,第一对腕极长。短蛸(O.ochellatus)腕短,各腕长度近等(图9-51A~B)。这两种我国沿海均有分布,短蛸多鲜食,肉嫩味美。长蛸干制品味美,鲜品为钩捕大型经济鱼类的饵料。船蛸(Argonauta)雄体小,无外壳。雌体大,背腕具翼状腺质膜,能分泌2次性石灰质壳。我国南海有分布(图9-51C)。
第三节 软体动物的经济意义
软体动物种类多,分布广,大多数种类与人类关系密切,有的可食用、入药或为家禽家畜的饲料等;不少有害的种类与吸虫病的传播有关。
食用的软体动物含丰富的蛋白质、无机盐和各种维生素,有的含大量糖原,且营养成分易溶于水,易消化吸收。淡水种类的田螺、蚬等,海产的鲍、泥螺、蚶、扇贝、牡蛎、乌贼、柔鱼等,均为习用食品,味鲜美,富营养。可入药的如鲍的壳(石决明)、乌贼内壳(海螵蛸)、海兔的卵群(海粉)等。利用帆蚌、珍珠贝等育珠,珍珠为高贵装饰品,又为工业原料,亦可药用。不少贝类的壳可制纽扣、螺钿等。许多螺类和双壳类为一些鱼类的天然饵料,淡水螺、蚌等可作家禽家畜的饲料,促进其生长发育,提高产量。因此许多种类已进行人工养殖。
有些双壳类如船蛆、海筍,钻木凿石而栖,危害海港建筑;附着生活的种类,可堵塞工业输水管道,造成严重损失;有些螺类如蜗牛、蛞蝓、玉螺等危害农作物或破坏贝类养殖;有的是海藻养殖的敌害,如锈凹螺(Chlorostoma)。不少淡水螺为吸虫的中间宿主,危害极大,如钉螺、沼螺等。
第四节 软体动物的系统发展
软体动物的海产种类个体发生中为螺旋型卵裂,且具有担轮幼虫,排泄器官为后肾管,这些特点均与环节动物尤其是多毛类近似。故有理由认为软体动物和环节动物在系统发生中有着共同的起源,在长期进化中,朝着不活动的生活方式发展,因而体节消失,产生了贝壳,运动器官和神经感官均趋于退化。
软体动物中单板纲、无板纲及多板纲较为原始,这几类的次生体腔发达,近似梯式神经;有的体呈蠕虫形,无壳,许多器官如鳃、肾、外壳等显示出分节排列现象。这些原始性状的存在,认为它们接近软体动物的原始祖先,各自独立发展一支。
腹足类较为原始,其生活方式活跃,头部发达。瓣鳃纲生活方式不活动,无头,但原始种类具楯鳃,足部具跖面,这与腹足纲接近。掘足纲头不明显,套膜在胚胎时为2片,后才愈合呈筒状,成对的肾,脑神经节与侧神经节分开,这些表明接近于原始的瓣鳃类。但掘足类无鳃,无心脏,贝壳筒形,又显示与其他纲动物在演化上较为疏远,可能是较早分出的一支。头足纲为一古老的类群,起源早,化石种类多。它们生殖腔与体腔相通,似无板纲;个体发生中,在胚胎早期无肾,似多板纲和无板纲;生殖导管来源于体腔导管,又似多板纲。由于头足类具有原始软体动物的特点,说明它们与软体动物的原始种类接近。但头足类有机结构复杂,神经系统高度集中,且为软骨质包围;眼的结构似脊椎动物;基本为闭管式循环系统;直接发生,无幼虫期。由于头足类既有原始性状,又有高度的进化特征,故推测它们可能很早分出的一支,沿着更为活跃的生活方式发展的一个独立的分支。
复习题
1.试述软体动物门的主要特征。
2.软体动物与环节动物在演化上有何亲缘关系,根据是什么?
3.软体动物分哪几纲,简述各纲的主要特征。
4.分析软体动物种类多、分布广与其形态结构和生活习性的关系。
5.分析比较多板类、腹足类、瓣鳃类及头足类的主要结构特点的异同。
6.软体动物有固着、穴居、爬行、游泳和寄生等不同生活方式,试述其特殊结构对生活方式的适应。
7.腹足纲分类(亚纲及目)的主要依据。
8.瓣鳃纲分类(目)的主要依据。
9.试述头足类对环境适应的结构特点。
10.了解软体动物与人类的利害关系。
本文标题:第九章 软体动物门(Mollusca)
手机页面:http://m.dljs.net/dlsk/dongwuxue/59424.html
本文地址:http://www.dljs.net/dlsk/dongwuxue/59424.html
