爬行纲在中生代曾盛极一时,种类和数量极其繁多,现存种类只包括鳄、龟、蜥蜴和蛇等动物。其中个体最大的是产于亚洲东南部的网蟒(Python reticulatus),全长可达9.8m,而最小的蜥蜴为斑点圆趾虎(Sphaerodactylus argus),全长却只有36mm。除南极地区外,分布几乎遍及全球而尤以南半球的种类更为繁多,能栖息于平原、山地、森林、草原、荒漠、海洋和内陆水域等各种生活环境,少数几种蜥蜴(西藏沙蜥Phrynocephalus theobaldi、红尾沙蜥Phrynocephalus eryth-rurus)的最高垂直分布点可达海拔五千多米的青藏高原寒漠地带。
第一节 爬行纲的主要特征
一、羊膜卵及其在动物演化史上的意义
在脊椎动物从水到陆的漫长进化历程中,动物体各系统器官在获得全面发展的基础上,由两栖动物产无羊膜卵转变成爬行动物产羊膜卵是一个极其重要的飞跃进步。羊膜卵的结构和发育特点,使羊膜动物彻底摆脱了它们在个体发育初期对水的依赖,这才可能确保脊椎动物在陆地上进行繁殖。
爬行动物产的羊膜卵为端黄卵,具有卵黄膜而缺乏适于水中发育的内胶膜和外胶膜,包裹在卵外的有输卵管壁所分泌和形成的蛋白、内外壳膜(shell membrane)和卵壳(shell)。卵壳坚韧,由石灰质或纤维质构成,能维持卵的形状、减少卵内水分蒸发、避免机械损伤和防止病原体侵入。卵壳表面有许多小孔,通气性良好,可保证胚胎发育时进行气体代谢。卵内有一个很大的卵黄囊,贮有丰富的卵黄,为发育期间的胚胎供给营养物质。
胚胎在发育期间,发生羊膜(amnion)、绒毛膜(chorion)和尿囊(allantois)等一系列胚膜是羊膜动物共有的特性,也是保证羊膜动物能在陆地上完成发育的重要适应。羊膜卵的胚胎发育到原肠期后,在胚体周围发生向上隆起的环状皱褶——羊膜绒毛膜褶,不断生长的环状皱褶由四周逐渐往中间聚拢,彼此愈合和打通后成为围绕着整个胚胎的2层膜,即内层的羊膜和外层的绒毛膜,两者之间是一个宽大的胚外体腔(exocoelom)。羊膜将胚胎包围在封闭的羊膜腔(amnioticcavity)内,腔内充满羊水,使胚胎悬浮于自身创造的一个水域环境中进行发育,能有效地防止干燥和各种外界损伤。绒毛膜紧贴于壳膜内面。胚胎在形成羊膜和绒毛膜的同时,还自消化道后部发生一个充当呼吸和排泄的器官,称为尿囊。尿囊位于胚外体腔内,外壁紧贴绒毛膜,因其表面和绒毛膜内壁上富有毛细血管,胚胎可通过多孔的壳膜和卵壳,同外界进行气体交换。此外,尿囊还作为一个容器盛纳胚胎新陈代谢所产生的尿酸(图18-1)。
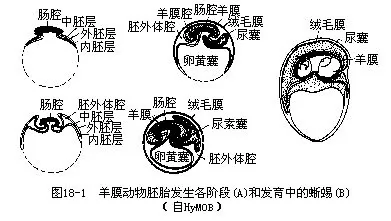
爬行动物获得产羊膜卵的特性后,毋需到水中繁殖,为爬行动物通过辐射适应向干旱地区分布及开拓新的生活环境创造了条件。
3亿年前,当具有高等和进步特征的新兴爬行动物在地球上出现后,很快就得到极大的发展,成为在地球上各种生态环境中占主导地位的动物。然而,爬行动物有机结构的完善程度并未达到动物界发展的顶峰,与更高等的鸟纲和兽纲比较,它们还有许多较为低级之处。因此爬行动物与脊椎动物中的无羊膜类一样,都属于变温动物,即它们自身活动产生的热量较少,以及体温调节机能的不完善,不足以维持恒定的体温,在很大程度上要受到环境的影响;它们不能生活于过低或过高的温度环境中,因而在严寒酷冷的冬季和炎热干旱的夏季里需要进行蛰眠。
二、爬行纲的躯体结构
(一)外形 爬行动物体表被有鳞片。身体外形差异显著,可分为基本形态的蜥蜴型(蜥蜴、鳄、楔齿蜥等)和特化形态的蛇型(蛇和蛇蜥等)、龟鳖型(龟和鳖等),分别适应于地栖、树栖、水栖和穴居等不同生活方式。除蛇型种类外,身体可明显地区分为头、颈、躯干和尾部,有活动性的眼睑(壁虎科种类例外),鼓膜下陷于外耳道的深处;四肢强健有力,前后肢均为五指(趾),末端具爪,善于攀爬、疾驰和挖掘活动;泄殖孔纵裂(鳄、龟)、横裂(蜥蜴、蛇)或圆形(龟、鳖);尾基较为粗大,往后逐渐变细,终于尾梢。
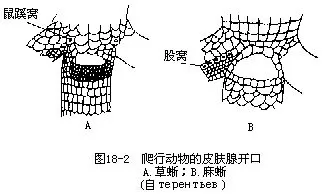
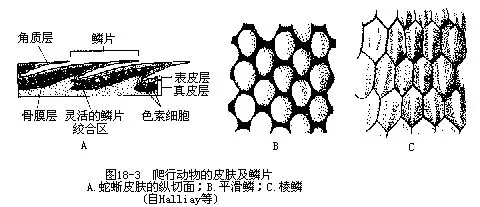
(二)皮肤 爬行动物的皮肤特点是表皮高度角质化,约有10层细胞构成,且外被角质鳞(horny scale),构成完整的鳞被,可以有效地防止体内水分的蒸发;角质鳞的彼此相邻部分角质层变薄,使鳞片稍能活动。树栖或善于攀爬的种类,体表常被发达的棱鳞(keeled scale)。鳄类在角质鳞下尚埋有真皮发生的骨板和腹膜肋(abdominal ribs);龟甲则由表皮形成的角质盾(hornyepidermal shield)与真皮来源的骨板(bony plate)共同愈合所成。皮肤干燥,皮肤腺很不发达,较常见的皮肤腺有位于草蜥、麻蜥、胎生蜥、腊皮蜥和壁虎科种类大腿内侧或泄殖孔前的股腺(fem-oral gland)。腺孔排列成行,称为鼠蹊窝(igunal pore)、股窝(femoral pore)和肛前窝(preanal pore)等,其分泌物于繁殖期常积贮在孔外,形成黄色的短刺状小突起,有助于交配时把持雌蜥(图18-2)。鳄的泄殖腔内及蛇尾的基部,以及龟类的颌下和胯间都具有一对臭腺(scent gland),它的分泌物有特殊气味,雄体能以此招诱异性或循着味源找到雌体进行逐偶交配。
爬行动物的鳞被有定期更换规律,称为蜕皮(ecdysis),然而,一生中的蜕皮次数及蜕皮方式因种而异。生长迅速的蛇类每两个月左右蜕皮一次,由于蛇的鳞片浮覆在真皮突起的顶部,又不与真皮紧密连接,因此在去旧换新的蜕皮过程中,在酶的作用下,将旧的表皮细胞层的基部溶解掉,使旧的表皮角质层与生发层细胞不断分裂所形成的新细胞层分离,把蛇皮完整地蜕下。
蜥蜴的蜕皮为成片脱落,不及蛇类明显;龟、鳄无定期蜕皮规律,通常是以不断更新替旧的方式进行的。
真皮由致密的纤维结缔组织构成,内含丰富的色素细胞(鳄的色素颗粒沉着在表皮角质层),由此组成色彩鲜艳的斑纹图案(图18-3)。许多蜥蜴(草蜥、捷蜥、石龙子、沙蜥等)的色素细胞在植物性神经或脑垂体等内分泌腺的调节下,控制其扩展和收缩,从而引起体色变化。避役类因善于快速变色而素有“变色龙”之称。蜥蜴的变色除了起到使身体与环境融为一色的保护作用外,还具有吸收地表幅射热及调温的功能。
(三)骨骼系统 爬行动物的骨骼系统大多由硬骨构成,骨骼的骨化程度高,很少保留软骨部分。
1.中轴骨骼
(1)颅骨较高而隆起,为高颅型(tropibasic type),表明颅腔扩展及脑容量已有明显增大。构成颅骨的软骨化骨和膜骨数目,在陆生脊椎动物中是最多的。
(2)有单枚枕髁,与第一枚颈椎关连。
(3)颅底由前颌骨和上颌骨的腭突、腭骨等的突起共同合成雏型的次生腭(secondary pala-ta),使口腔中的内鼻孔位置后移。次生腭的结构在鳄类中尤为发达,是适应于水中捕食的特化现象。(图18-4)。

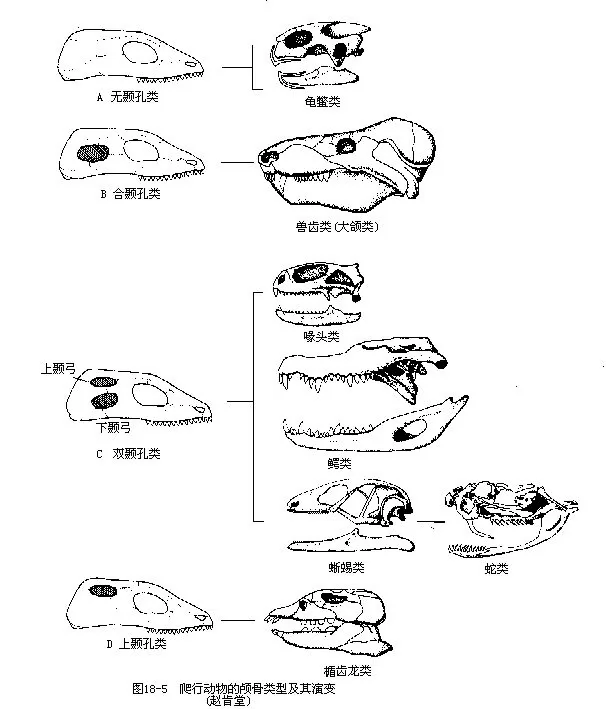
(4)颅骨两侧在眼眶后方出现1~2个颞孔(temporal fossa),颞孔是由于原来附生咬肌的颞部向内低陷所形成的孔洞,当咬肌等收缩时,可使膨大的肌腹部向颞孔凸出而纳入其间。通常,颞孔位于眶后骨、颧骨、鳞骨、方轭骨和顶骨之间,颞孔上、下的骨弓分别称为上颞弓和下颞弓。根据颅骨上颞孔的有无及孔的位置,可将爬行动物分为无颞孔类(anapsida)、上颞孔类(parapsida)、合颞孔类(synap-sida)和双颞孔类(diapsida)等4个类型。无颞孔类的颅骨无颞孔及颞弓,是古爬行动物的原始颅骨类型;现存海龟类也缺乏颞孔,次生性的变化,上颞孔类、合颞孔类和双颞孔类均由无颞孔类演变而来。上颞孔类的颅骨只有单个上颞孔,上颞弓由眶后骨和鳞骨构成,具有这种类型颅骨的爬行动物是鱼龙类(Ichthyosauria)和合颞孔类的颅骨一侧各有单个颞孔,被眶后骨、鳞骨和颧骨所围,以眶后骨和鳞骨构成上颞弓,古爬行动物中的兽齿类(Theriodontia)属之,现代哺乳纲动物即由此演进而来。双颞孔类的颅骨两侧具有上、下2个颞孔,现存的鳄、蜥蜴、蛇等爬行动物的颅骨均属本类,然而在进化过程中都发生了不同程度的特化,只有楔齿蜥(Sphenodon)和鳄类还保留着典型的双颞孔类的颅骨。蜥蜴类因方轭骨萎缩或退化,不能构成下颞孔的下颞弓,颅侧仅留下单个上颞孔;蛇类在蜥蜴退化方轭骨的基础上,又进一步失去眶后骨及颧骨,从而完全消失了作为眼和上、下颞孔分界的骨弓,使三者合成一个大孔(图18-5)。
(5)很多种类在两眼窝之间具有薄骨片形成的眶间隔(interorbital septum)。
(6)下颌除了关节骨外,还有齿骨、夹板骨(splenial)、隅骨(angular)、上隅骨(surpra angular)和冠状骨(Coronoid)等多块膜成骨参与组成。
(7)脊柱已分化成陆栖脊椎动物共有的颈椎、胸椎、腰椎、荐椎、尾椎等5个区域。出现了2枚荐椎,第一、第二颈椎分别特化为寰椎(atlas)和枢椎(axis)。荐椎数目的增多及其与腰带的牢固连接,加强了后肢承受体重负荷。寰椎前部与颅骨的枕髁关连,枢椎的齿突伸入寰椎,构成可动联结,使头部获得更大的灵活性,从而使头部既能上下运动,又能转动(图18-6)。

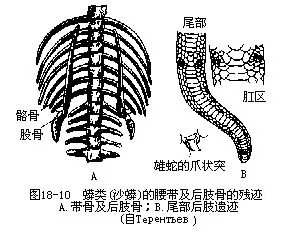
有些蜥蜴(蛇蜥、麻蜥、草蜥、石龙子、蝘蜒、壁虎等)的尾椎中部有一个能引起断尾行为的自残部位,这是尾椎骨在形成过程中前、后两半部未曾愈合而特化的结构。一旦蜥尾遭受拉、压、挤等机械刺激时,附生在自残部位前、后的尾肌分别往不同方向作强烈的不协调收缩,于是就会在尾椎骨的某个自残部位处断裂,连同肌肉和皮肤一起发生自残(autotomy)断尾现象。由于自残部位的细胞始终保持着增殖分化能力,因此残尾断面可重新长出再生尾(图18-7)。

颈椎、胸椎和腰椎两侧都附生发达的肋骨,每根肋骨一般由背段的硬骨和腹段的软骨合成,除龟鳖类和蛇类外,爬行动物前面一部分胸椎的肋骨均与腹中线的胸骨(sternum)连接成胸廓(throax)。胸廓为羊膜动物所特有,是与保护内脏器官和加强呼吸作用的机能密切相关的,同时也为前肢肌肉提供了附着点;肋骨附有肋间肌,它们的收缩可造成胸廓有节奏性的扩展和缩小,协同呼吸运动的完成。鳄类和楔齿蜥于胸骨后方有浮生在皮下的腹膜肋,是来源于真皮的退化骨板(图18-8)。
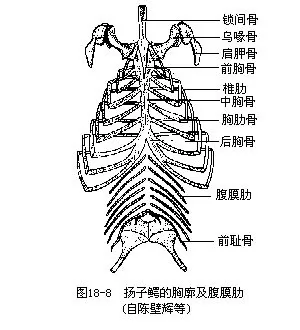
2.附肢骨骼
(1)肩带内有一块十字形的上胸骨,或称锁间骨(interclavicle),该骨于龟鳖类中转化为腹甲的内板(entoplastron)。
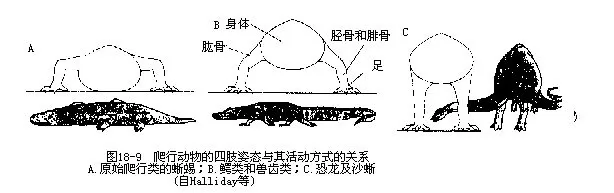
(2)四肢与身体的长轴呈横出的直角相交,且其肩臼浅小,因此不少爬行动物在停息或爬动时,都保持着腹部贴地的姿态,只有沙蜥等能使四肢肘(elbow)或膝(knee)部以下的部分,转向腹部下方,将蜥体抬离地面疾驰奔跑(图18-9)。后肢的踵关节位于两列跗骨之间,这种关节方式称为跗间关节(intratarsal joint)。蛇蜥类(Ophisaurus)四肢消失,但仍留有带骨;蛇类既无四肢又无带骨,然而蟒科、盲蛇科(Typhlopidae)和瘦盲蛇科(Leptotyphopidae)动物的体内,则留有退化的髂骨和股骨,而泄殖孔两侧还有一对角质的爪状突,也是后肢残迹的一部分(图18-10)。
(四)肌肉系统
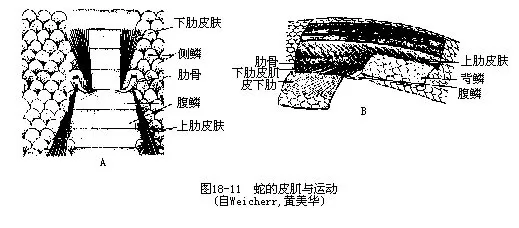
1.皮肌(skin muscle)一般起自躯干肌、附肢肌或咽部肌肉而止于皮肤。蛇是爬行动物中皮肌最发达的类群,皮肌收缩可引起皮肤及其附属的鳞片产生活动。骨具有较大的活动性,其远端和中段各附生着两对肋皮肌(cotocutaneous),分别与紧贴皮肌的前、后腹鳞相连。收缩时能使腹鳞运动凭藉反作用的推力,使蛇体蜿蜒爬行前进(图19-11)。此外,蜥蜴和龟的颈括约肌也属于皮肌。
2.咬肌始于颞部及上颌后部而止点位于下颌的颞肌(temporalis)和咬肌(masseter),均为闭口肌,起止于舌弓及下颌骨腹面的二腹肌为开口肌,是前两块肌肉的拮抗肌。咬肌因颞孔的出现,收缩时肌腹可以容纳在窝内,使咬啮机能得到了加强而变得有力。翼肌使下颌可作前后左右各个方向的运动,增强了捕食能力。
3.肋间肌(intercostal muscle)是营胸腹式呼吸的陆栖脊椎动物的特有肌肉。肋间肌位于胸部表层肋上肌下方的相邻两枚肋骨之间,可分为外肋间肌和内肋间肌,用于调节肋骨升降,控制胸腹腔的体积变化,并协同腹壁肌肉完成呼吸作用(图18-12)。
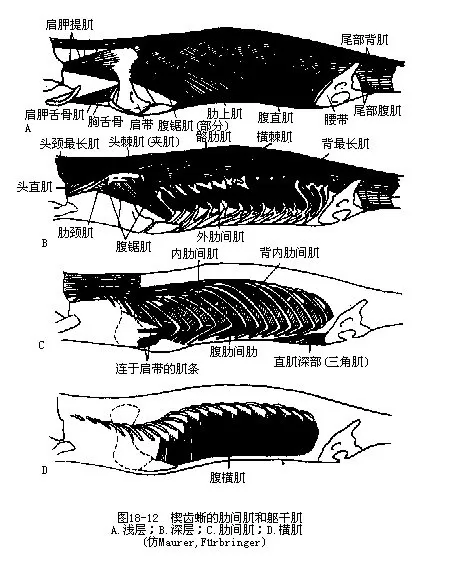
4.躯干肌 躯干肌因四肢发达而渐趋萎缩。背部的主要肌肉是最长肌,担负着脊柱上、下屈曲的机能。背肌在两侧还分化出一层薄片肌,为髂肋肌(iliocostales),往下伸展到腹壁,止于肋骨侧面。背最长肌和髂肋肌均起自颅骨枕区后缘,肌肉收缩与头、颈部的转动有关。腹肌由表及里仍为外斜肌、内斜肌、横肌构成,腹直肌也发育良好。
5.附肢肌四肢上部的肌肉粗大,前臂肌大多起自背部、体侧、肩带(如背阔饥三角肌和三头肌等),收缩时可前举和伸展前肢;后肢有位于腰腿之间的耻坐股肌、髂胫肌,腿部的股胫肌和臀部肌肉等,主要机能是把大腿拉向内侧和使膝关节闭合,将动物体抬离地面并往前爬动。
(五)消化系统口腔与咽有明显分界,有发达的唇腺(labialgland)、腭腺(palatinegland)、舌腺(lingual gland)和舌下腺(sublingual gland)等口腔腺,其分泌物有助于湿润食物和吞咽动作的完成。舌还具有示警器(蓝尾石龙子)、感觉器(蛇、蜥蜴)或捕食器(避役)的作用。鳄舌厚而宽,龟的舌形短宽,舌面有敏感的触觉,但都粘连于口底不能伸出口外。蜥蜴类的舌大多平扁而圆,有伸缩性。蛇舌形甚细长,缩藏在舌鞘内,舌上缺少味蕾,故无味觉作用,但能把空气中的气体分子溶附在舌上,并通过特有的犁鼻器感知嗅觉。避役类的舌,内为纵肌,外围环肌,顶端膨大而富粘性,平时舌压缩在口中的鞘套里,捕食时因舌内快速充血,环肌强烈收缩,将舌头从口中直射出去,在它射向昆虫等捕捉对象的瞬间,可以达到与蜥体等同的长度,并能准确无误地粘住猎物。
除龟鳖类的颌上无齿而颌缘覆有角鞘外,其他爬行动物的两颌分别长有各种型式的牙齿,受损脱落后可不断更新长出再生牙。通常按其着生位置不同,可分端生齿(如飞蜥、沙蜥)、侧生齿(大多数蜥蜴及蛇)、和槽生齿(如鳄类)(图18-13)。爬行动物的牙齿主要着生在上、下颌,但是也可以出现在蛇类的犁骨和翼骨上;楔齿蜥的幼体两颌及犁骨、翼骨都有牙齿,这种情形与两栖动物一致而在爬行动物中却是绝无仅有的。大多数种类的牙为同型齿(homodont),只有鳄类和少数鬣蜥科蜥蜴(沙蜥)有初步分化为异型齿的趋势。毒牙(fangs)是毒蛇前颌骨和上颌骨上的少数几枚特化的大牙,因表面有沟或中央有管而分别称为沟牙(grooved tooth)及管牙(canaliculated tooth),通过毒牙基部的排毒导管与毒腺(toxic gland)相联。沟牙因着生的位置不同又有前沟牙和后沟牙之分。毒腺由上唇腺转化而来,一般位于眼后部、口角上方或上颌外侧,毒腺外覆有坚韧的结缔组织,内侧具压毒腺肌,有压挤毒腺及牵引启闭下颌的作用。毒蛇咬物的同时,可通过肌肉对毒腺的压挤,迫使其中的毒液输入排毒导管,经毒牙的沟、管排出,注入捕获物体内(图18-14)。


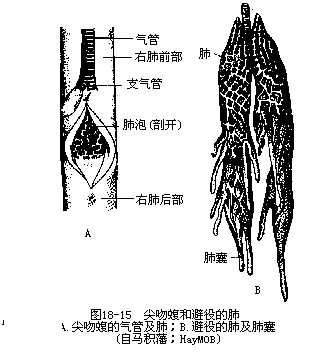
(六)呼吸系统内鼻孔呈裂缝状,开口在腭顶的中部或咽喉附近。随着颈的分化,呼吸道出现了管壁有软骨环所支持的气管(trachea),并由其后端分成一对支气管(bronchi)分别通入左、右肺脏内。肺脏是一对位于胸腹腔前部背面的呼吸器官,形似囊状,内部具有复杂的间隔,使之分隔成无数蜂窝状小室,并分布着极其丰富的肺动脉和肺静脉的微血管,因而比两栖动物能更有效地扩大与空气接触及交换气体的表面积。在蛇类和蛇蜥等无肢爬行动物中,肺脏的位置常出现两侧互不对称现象,左肺大多萎缩或退化(眼镜蛇科、海蛇科和一部分蝰蛇科种类),成为失去呼吸机能的残留器官。蝮蛇和避役类的肺脏结构比较特殊,前部为呼吸部,后部内壁平滑并伸出若干膨大的气囊,分布于内脏间,称为贮气部,无交换气体作用,这种结构在进化到鸟类时获得更完善的发展(图18-15)。
爬行动物除了像两栖动物可借助口底运动进行口咽式呼吸外,同时还发展了羊膜动物共有的胸腹式呼吸。胸腹式呼吸依靠外肋间肌收缩,提起肋骨,扩展胸腔,吸入空气进肺,当内肋间肌收缩时,可牵引肋骨后降,胸腔缩小,空气从肺内呼出,呼吸作用就是通过胸腔有节奏地扩张和缩小的过程完成气体交换的。
水栖龟鳖类能较长时间潜伏于水底,此时主要以咽壁和突出在泄殖腔两侧的副膀胱(或称肛囊)作为辅助呼吸器官。爬行动物的喉头与无尾两栖类近似,绝大多数种类的喉门都有一对上皮粘膜褶襞,但不形成声带,所以无法发声。壁虎科和避役科的蜥蜴在喉门有声带,能产生高低不等的喉音(guttural noises),其中尤以大壁虎发出的“蛤—蚧”之声为人们所熟悉。
(七)循环系统 爬行动物的血液为不完全双循环,主要特点是:
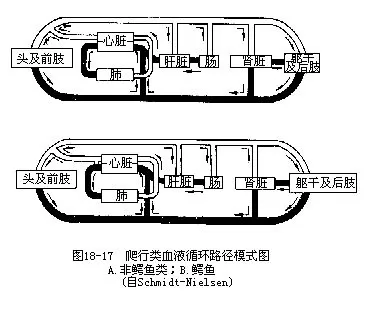
1.心脏包括两心房、一心室,静脉窦不发达,一部分被并入右心房,动脉圆锥已退化消失。蜥蜴类、蛇类和龟鳖类由于心室腹壁出现室间隔,使心脏形成分隔不完全的四室,在心脏收缩时,室间隔可抵达心室背壁,在短时间内使心室隔开成左、右两半,因而多氧血和少氧血的分流循环比两栖类益趋完善。鳄类具有完整的室间隔,仅在左、右体动脉的基部留有一个沟通2个心室的潘氏孔(foramen of Panizzae)(图18-16)。

2.离心的血液分别由左、右心室导出,流入3对动脉弓,其中肺动脉弓和左体动脉弓出自心室右侧和中央部,右体动脉弓则从心室左侧发出。心室在室间隔不完全处形成一个静脉腔,多氧血由肺静脉返回左心房、左心室,并有部分血液通过静脉腔流入右心室内,所以从右心室中部导出的左体动脉弓内,血液中的含氧量与左心室中的血液基本一致,或只混有极少量的少氧血。实际上仅右心室的右侧才是真正的少氧血。
3.通入头部的颈总动脉由右体动脉弓发出。
4.右体动脉弓远较左体动脉弓粗大,后者始于心室的右中部,其中的血液主要来自静脉腔,血液也为多氧血,因此左、右体动脉弓合成的背大动脉中,混合血的成分极少。
5.静脉系统近似两栖动物,返回心脏的主要静脉包括一对前大静脉、一条后大静脉、一条肝门静脉和一对肾门静脉。肾静脉逐渐趋于退化,后肢流向心脏的血液只有一部分在进入肾脏时分散为毛细血管,构成肾门静脉,另一部分则穿越肾脏直接汇入后大静脉(图18-17,图18-18)。

(八)排泄系统爬行动物与所有羊膜动物一样,以后肾(metanephros)作为滤尿器官。肾脏紧贴于身体后半部的背壁,左右各一,其位置在鳄类和蛇类中并不完全对称,右肾较左肾稍前。肾脏的体积因种类而异,但大多呈分叶状,爬行动物肾脏的基本结构和功能,同两栖动物并无本质区别,但是肾脏内的肾单位数目却远远多于后者,泌尿能力大为增强,并通过专用导管输送尿液,在输精管或输卵管开口的前内侧进入泄殖腔(图18-19)。

爬行动物除鳄类和蛇类外,泄殖腔的腹面均有膀胱,羊膜动物的膀胱由胚胎期的尿囊基部扩大形成,故称尿囊膀胱(allantoic bladder)。大多数爬行动物泌排的尿液中,其含氮废物主要是尿酸(uric ac-id)和尿酸盐,它们比尿素(urea)难溶于水,所以常在尿液中沉淀成白色半固态物质。这种物质的成分很复杂,包括钠、钾以及尿酸的氨盐,其中的钠、钾离子常以尿酸盐(urate salt)的形式,通过泄殖腔随粪便排出,而水分在这些物质沉淀时,又被输尿导管、大肠和膀胱重新吸收进入血液内,用于再产生尿和沉淀。这种重复周转对于干旱地区生活的爬行动物减少体液丧失和保持肾内不致形成高于血浆的渗透压,都具有十分重要的适应意义。
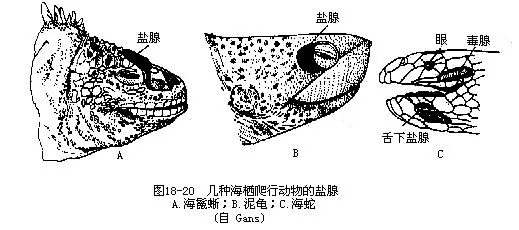
海龟、海鬣蜥(Amblyrhnchus cristatus)、海蛇和有些鳄(扬子鳄、湾鳄Crocodylus porosus、美洲鳄Crocodylus acutus、密河鳄Alligator mississippiensis、眼镜鳄 Caimen crocodilus等)具有特殊的排盐器官——盐腺(salt gland),执行肾外盐排泄(extra renal salt excretion)机能,通过盐腺分泌物将血液中多余的盐分带出体外。盐腺的位置视动物种类而不同,海龟位于眼后上方;海鬣蜥位于鼻部嗅囊外,分泌的含盐液体经小管流入鼻道,并形成氯化钠或氯化钾的结晶;海蛇的盐腺位于舌下;鳄类的舌腺位于舌中部及其两侧,扬子鳄的舌腺既有单管腺,也有复杂的泡沫状腺,数量约100个,兼有泌盐和分泌粘液的功能(图18-20)。有人认为在某些爬行动物中,盐腺的重要性甚至超过肾脏,对于体内盐水平衡和酸碱平衡均有重要意义。
爬行动物在胚胎发育期间,羊膜卵的结构不但能保护胚体和有效地阻止卵内的失水,而且还能以较小的体积来盛纳通过尿囊所排出的尿酸盐等代谢废物。

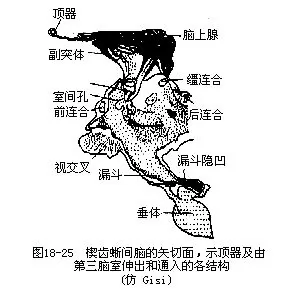
(九)神经系统爬行动物脑的各部已经不是完全排列在同一平面上,延脑发展出颈曲。大脑半球的体积明显地超过其他各部脑,然而增大和加厚的主要部分仍局限于大脑底部的纹状体,不过它的重要性已经足以同中脑视叶相提并论了;大脑半球的顶壁及其两侧基本上还是原脑皮,但是开始出现锥体细胞(pyramidal cell),并聚集成神经细胞层,构成大脑表层的新脑皮(neopalli-um)。新脑皮在爬行动物中尚处在系统发生的早期阶段。间脑在背面几乎难于辨识,由此发出细小的脑上腺(epiphysis)和顶器(parietal organ)。中脑背面为一对圆形的视叶,依然是爬行动物的高级神经中枢,由视神经传入至下丘脑的神经纤维中,大部分都直接通到视叶,但是也有少数神经纤维经丘脑达于大脑(图18-21),这表明从爬行动物起神经活动已有向大脑逐渐集中的趋势,发展到哺乳类则达到了高峰。中脑在蟒蛇和响尾蛇已分化为四叠体(corpora quadrigemina),通常前面一对视叶的体积较小。小脑比两栖动物大而不及鱼类发达,这与爬行动物缓慢活动的生活习性有关。善于游泳的水生爬行动物(龟鳖类)小脑发育良好,其后缘一般都覆盖着延脑菱形窝的前半部,而鳄类的小脑两侧甚至还开始分化出一对小形的小脑耳,是鸟、兽中蚓体和小脑鬈的前身,其机能与平衡相关(图18-22)。
爬行动物开始具有12对脑神经,鳄和龟鳖类的脊副神经(spinal accessory nerve)已经从迷走神经分化出来,并被包围在颅骨内。蛇和蜥蜴仅有脑神经11对,蛇的迷走神经和脊副神经尚未分离,而鳄蜥的脊副神经离脑后即并入迷走神经,未成独立的游离神经。

(十)感觉器官
1.嗅觉 鼻甲骨(conchae)是爬行动物鼻腔内首次出现的结构,并在鳄类中已发展到具有3个鼻甲骨的水平。鼻甲骨的表面覆有嗅上皮,分布着嗅神经和嗅觉感觉细胞,是真正的嗅觉区。龟鳖类的鼻甲骨不发达。
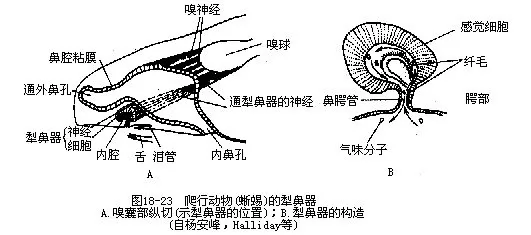
蜥蜴和蛇类的犁鼻器(Jacobson's organ)十分发达,以独立的鼻腭管开口在口腔顶部。犁鼻器的内壁覆有感觉上皮和嗅粘膜,并通过位于感觉上皮深层的犁鼻神经(嗅神经的分支)与脑相连,司嗅觉。由于犁鼻器不与外界直接相通,因此它的这部分嗅觉作用主要靠舌的活动才得以完成。蜥蜴和蛇的舌尖经常从上唇正中的缝隙内频繁地伸出口外活动,旨在探知和搜集空气中的各种化学气味物质,当舌尖缩回口腔时,即进入犁鼻器将化学分子溶解于嗅粘膜上,产生嗅觉(图18-23)。鳄类无犁鼻器,龟鳖类虽有犁鼻器,但只突入鼻腔,并不象蜥蜴那样通入口腔。
2.视觉 除了蛇和蜥蜴类中的蛇蜥、壁虎科动物外,爬行动物都有活动性的上、下眼睑和瞬膜,在龟鳖类、鳄类和蜥蜴中出现了泪腺(lachrymal gland),其分泌物经鼻泪管由鼻腔排出。蛇眼表面盖有一层透明的薄膜,是由上、下眼睑扩展、愈合和特化而成,有保护眼球的作用,蜕皮时可随同全身的角质鳞一起完整地蜕掉。
眼球的构造比较典型,视觉的调节是依靠改变晶体与视网膜之间的距离以及改变晶体的形状来控制的。晶体略成扁圆形,在正常情况下适于远视,其周围的“赤道”部连接着悬挂晶体的睫状体(ciliary body),内有横纹肌性质的睫状肌(ciliary muscle),不但能移动晶体的前后位置,调节视距,收缩时还能改变晶体的凸度,使之成为圆形适应近视。蛇眼内的睫状肌退化,使晶体变形的视距调节作用,由虹膜括约肌的收缩代为执行。
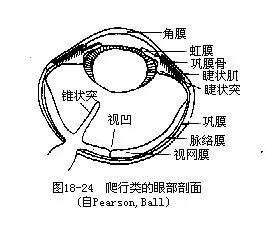
爬行动物眼球的后眼房内通常有一个从脉络膜突出的锥状突(conus papillaris),主要由结缔组织构成,内具丰富的毛细血管,有营养眼球的功能。眼球的巩膜中有一圈呈覆瓦状排列的环形小骨片,为巩膜骨(scleral ossicle),这是某些古爬行动物遗留下来的性状,也是与现代鸟类所共有的特征,有保护眼球的作用(图18-24 )。
楔齿蜥和一些(鬣蜥科、蜥蜴科)蜥蜴于两眼稍后方的头部正中,有单个顶眼(parietal eye),顶眼位于间脑顶器的顶端,埋于皮下,通过顶间鳞上的颅顶孔隐约可见,光线也能由颅顶孔透入。顶眼的基本结构与正常眼相似,具有很小的角膜、水晶体和视网膜,但是不能使物体在视网膜上成像,仅有感光功能。顶眼对于蜥蜴调节和适度地利用日光热能起着极其重要的作用,还与蜥蜴某些周期性的生命活动和识别归途具有密切关系。爬行动物的顶眼来源于旁松果体(parapinealbody),但是七鳃鳗的顶眼却来自松果体(pineal body=脑上腺epiphysis),有人认为最原始的脊椎动物可能存在2个顶眼,而其排列方式则是一前一后,并非左右对称,有些盾皮鱼类头骨化石的结构,可以初步证实这一推论的正确。
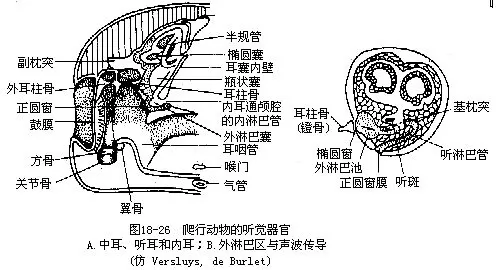
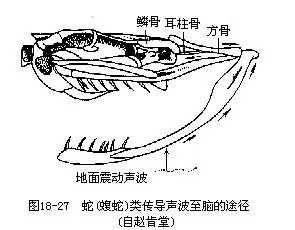
3.听觉 爬行动物的听觉器官由于鼓膜下陷而出现了雏形的外耳(听)道,中耳内有耳柱骨和耳咽管,耳柱骨的外、内两端分别接触鼓膜和紧贴内耳的椭圆窗(fenestra ovalis),椭圆窗之下还出现了前所未有的正圆窗(fenestra rotunda),窗外盖有薄膜,使内耳中淋巴液的流动有了回旋余地。内耳的膜迷路构造与两栖动物大致相同,但是司听觉的瓶状体有了明显扩大,瓶状体的基部是一个由感觉毛细胞组成、并被胶质基膜掩盖的基乳突,为爬行动物的重要听觉区。声波通过鼓膜、听骨的振动,自椭圆窗传入内耳,再由一个外淋巴管绕过瓶状体的基乳突抵达正圆窗。外淋巴管与基乳突下的感觉毛细胞仅以一薄层的基膜相隔,基膜的振动可激发起感觉毛细胞的同步振动,并由毛细胞下方的神经传入脑内感知听觉。鳄类的瓶状体延长成卷曲的耳蜗(cochlea),内有螺旋器,是真正的听觉器,雌雄鳄在繁殖期相互之间以吼声传递讯息,也可证明其听觉器官发达(图18-26)。蛇类适应穴居生活,中耳、鼓膜和耳咽管均已退化,但是耳柱骨仍保留,它能由下颌骨敏锐地接受从地面振动传来的声波,并通过颅骨上的方骨及与之关连的耳柱骨传入内耳,从而感受听觉(图18-27)。
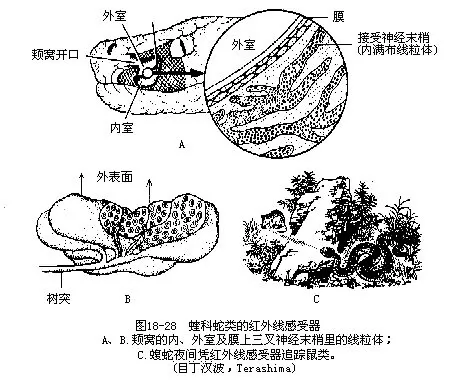
4.红外线感受器(infrared receptor)这是蝰科蝮亚科和蟒科蛇类头部特有的热能感受器,即位于蝮蛇、竹叶青、响尾蛇等眼鼻之间的颊窝(facial pit)及蟒类唇鳞表面的唇窝(labial pit)。尖吻蝮的颊窝是一对三角形凹陷,窝内有一层厚仅10μm~15μm的薄膜,将窝分隔为内、外两室,外室较大,开口朝外;内室位于颊窝深处,以一细管导向眼前角,借一小孔通至皮肤表面,可调节内、外室之间的压力,孔口有括约肌。颊窝膜的内壁上密布着三叉神经分支的神经末梢,其终端略为膨大,内部充满线粒体(mitochondrion)。这是一种极其敏感的热能检测器,只需123.68×10-5J/cm2的微弱能量就能使之激活,感知在一定范围内的其他动物,并在35ms的瞬息之间迅速作出反应,其效能为现今最灵敏的红外线探测器所不及。人们可以在观察颊窝膜上的神经末梢时,见到其线粒体的形态发生热缩冷胀的相应改变,这就表明热能感受器的作用与线粒体的关系殊为密切。蛇的颊窝不但能觉察到周围环境中的温血动物,甚至还能确定该动物的位置,从而进行追踪和准备袭击,因此也可视为一种有助于夜间觅食的特殊感觉器官(图18-28)。唇窝是位于蟒类吻鳞或上唇鳞表面的小形洼陷。其结构与蝮亚科蛇类的颊窝相似,也是一种热测位器,能感知0.026℃的温差变化,灵敏度稍逊于颊窝。
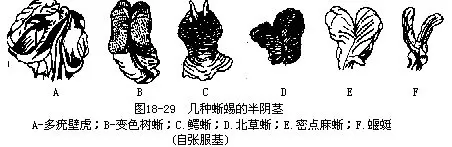
(十一)生殖系统 爬行动物营体内受精,除产于新西兰的楔齿蜥外,雄性均有交配器。蛇和蜥蜴的雄性交配器是一对半阴茎(hemipenis),平时不显露体外,埋藏在泄殖腔后方而位居尾基腹面的2个肌质的阴茎囊中,故有些种类的雄蜥尾基部常显得比较膨大;阴茎囊由薄层的环肌构成,收缩时能压挤半阴茎而使之翻出泄殖腔外,囊的末端藉韧带联接在尾肌上。半阴茎的腹面正中有深凹的精沟,并由其前方分叉至顶端的龟头;交配时,2个半阴茎同时从泄殖腔内翻出,但是只有一侧的半阴茎插入雌体泄殖腔中;半阴茎的基部与泄殖腔相通,可将顺输精管而下的精液,沿着半阴茎的精沟,注入雌性体内。各种动物的半阴茎形态不一,可作为分类及探索动物间亲缘关系的依据(图18-29)。
雄性龟、鳖的泄殖腔腹面有单个交配器,称为阴茎(penis),内有海绵体,能充血勃起伸出体外交配(图18-30)。
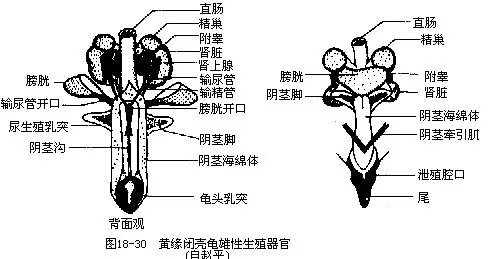
雌性有卵巢和输卵管各一对。输卵管上段为开口于体腔的喇叭口,中段因有蛋白腺而称为蛋白分泌部(蜥蜴和蛇类无蛋白腺),下段是能分泌形成革质(蜥蜴和蛇类)或石灰质(龟、鳖、鳄)卵壳的壳腺部,输卵管末端开口于泄殖腔,雌性的龟和鳖在泄殖腔壁上有一个不甚明显的阴蒂(clitoris),是与雄体阴茎同源器官的结构。
绝大多数爬行动物以卵生方式繁殖,主要依靠阳光的温度或植物腐败发酵产生的热量进行孵化。
有些古北界高纬度高海拔地区的蛇和蜥蜴,以及海蛇类营卵胎生(ovoviviparity)。受精卵在输卵管内发育成仔体时产出。少数种类(石龙子 Eumeces chinensis、蓝尾石龙子Eumeces ele-gans)的受精卵在母体的输卵管内已初步发育,至产卵前可进入器官形成阶段,并出现了脑泡及眼点等,是介于卵生和卵胎生之间的一种过渡类型,可称为亚卵胎生。迄今所知,我国至少有12种蜥蜴和25种蛇营胎生或卵胎生,约占蜥蜴总数的7.8%,占全部蛇种的13.7%,常见种类有南疆沙蜥、蝘蜓(Lygosoma indica)、秦岭滑蜥、密点麻蜥、鳄蜥、大盲蛇、沙蟒、红点锦蛇、水赤链蛇、水蛇、草原蝰、蝮蛇、白唇竹叶青、海蛇、小头海蛇和海蝰蛇等。
第二节 爬行纲分类
现存的爬行动物约有6550多种,我国约有380多种,除喙头蜥目外,其余4目国内均有分布。
一、喙头蜥目(Rhynchocephaliformes)
本目是爬行动物中最古老的类群之一,它们大多生活在下二叠纪和三叠纪,曾经广泛分布于全球南北的欧、亚、非和拉丁美洲,现在只存楔齿蜥(Sphenodon punctatum)1种,见于新西兰北方柯克海峡的一些小海岛上,总共不足千尾,已濒临绝灭的边缘。由于楔齿蜥具有一系列类似古爬行动物的原始特征,因此在动物学上有“活化石”之称。
楔齿蜥体形似蜥,成体头部前端呈鸟喙状,口内无齿,故又名喙头蜥。体长50cm~70cm,体被小形粒鳞,头、颈和背脊中央具一纵列锯齿状鬣鳞。周身黑绿色,体侧缀有白色斑点。楔齿蜥的原始特征有:双凹型椎骨,椎体间仍留有脊索的残余;腹部的肌肉中有腹膜肋;雄性无交配器;头骨为双颞孔型,有两个完整的颞孔和上、下颞弓;方骨不能活动;端生齿;顶眼十分发达,并有晶体、角膜、视网膜等正常眼的结构。
楔齿蜥与海鸟有“鸟蜥共栖”现象,它们昼伏夜出,白天居住在黑鹱、信天翁、海燕、海鸬鹚的洞穴内,夜间外出觅食昆虫、蠕虫和鱼虾等(图18-31)。11月至翌年2月共产卵8~15枚,孵化期约13个月。幼体生长发育很慢,20年左右才性成熟,曾有生活77年的记录,据称最长寿命可达百年。

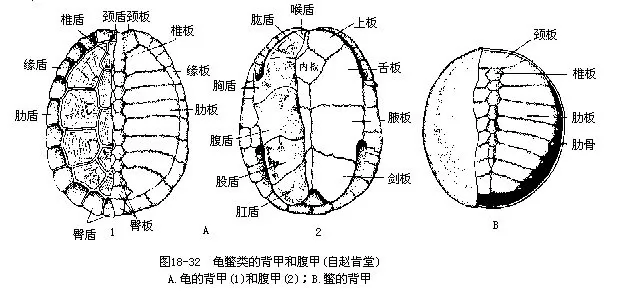
二、龟鳖目(Testudoformes或Chelonia)
龟鳖目是爬行动物中的特化类群。身体宽短,躯干部被包含在坚固的骨质硬壳内,头、颈、四肢和尾外露,但大多数种类可缩入壳中。硬壳由背甲和腹甲组成,外覆角质板或软皮。胸腰部的椎骨连同肋骨一起与背甲互相愈合;肩带位于肋骨的腹面,这是脊椎动物中绝无仅有的现象;无胸骨,也不形成完整的胸廓,上胸骨和锁骨分别参与了腹甲(内板和上板)的组成。颅骨上没有颞孔;方骨与颅骨团结,不能活动;颌和口内无齿,代之以颌缘的角质鞘;舌无伸缩性;有瞬膜和活动性眼睑。泄殖孔纵裂或圆形。雄性具单个交配器。卵壳钙质或革质(海龟),每年产卵2~3次,一次的产卵数为几十个至200枚以上。龟鳖的寿命较长,一般可活数十年,据报道巨龟(Testudogigantea)和海龟的最长寿命可达150~250年。现存种类有250多种,分属于13科,大多分布于温带和热带地区,我国产37种。代表性的种类有(图18-33):
(一)平胸龟科(Platysternidae)本科仅1属1种,即平胸龟(Platysternon megacephalum)。头大颌强,上喙钩曲呈鹰嘴状,俗称大头龟或鹰嘴龟;尾长,约与腹甲长相等;头、肢及尾不能缩入龟壳内;龟壳扁平,背、腹甲以韧带连接。生活于山区溪流中,夜间活动,能凭藉钩喙、利爪和长尾的帮助,越过障碍物或攀援上树。分布于东南亚地区和我国长江以南诸省。
(二)龟科(Testudinidae)头顶或至少头背前部被鳞,颅骨颞区有凹陷。龟壳坚硬,绝大多数种类的背甲和腹甲在甲桥处以骨缝接合,但闭壳龟(Cistoclemmys)和摄龟(Cyclemys)则借韧带相连;龟壳的骨板外覆以表皮性角盾。头、颈、四肢及尾均可缩入壳内。附肢粗壮,爪钝而强,指、趾间有蹼或无蹼,约有90多种,除大洋洲外,广泛分布于世界各地。具有代表性的种类有:
象龟(Geochelone elephantopus)背甲隆凸,高达50cm以上,全长140cm,体重约 250余kg,是世界上最大的陆生龟类。分布于南太平洋厄瓜多尔科隆群岛。
四爪陆龟(Testudo horsfieldi)头顶有对称大鳞,吻短;背甲半球形而高;四肢圆柱状,4爪,指、趾间无蹼;尾短,末端有一角质的爪状结节。栖于我国新疆霍城地区的沙漠中,每年4~7月为活动季节,挖洞隐居,白昼外出,其余诸月均深埋沙穴,处于蛰眠状态,有旱龟之称。
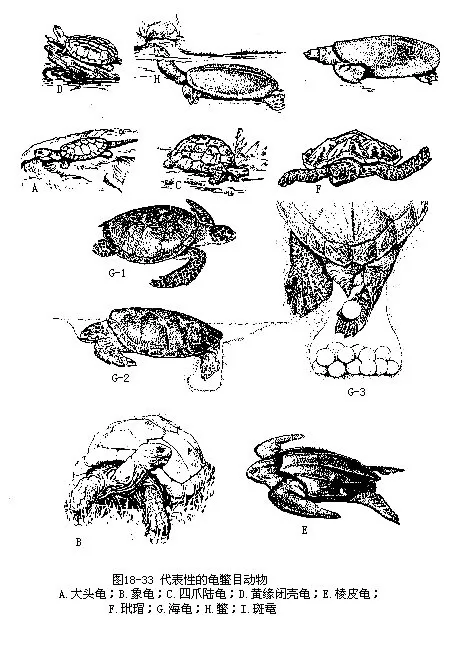
乌龟(Chinemys reevesii)又名金龟和草龟,是我国最常见的龟类,除西北地区和青藏高原外,其它各省都有分布。头顶前部平滑,后方被粒鳞,喙缘的角质较弱;背甲有3条纵棱;指、趾间全蹼。生活于江河、湖沼和池塘内,在向阳有荫的岸边松土处挖洞产卵。杂食性。
黄缘闭壳龟(Cistoclemmys flavomarginata)头顶皮肤光滑,鼓膜圆而清晰,上颌钩状。背甲的脊棱明显;腹甲的胸盾和腹盾与其相应的骨板之间以韧带相连,使腹甲的前、后两半可以活动,在头、尾及四肢缩入壳内时,完全闭合于背甲,闭壳龟的名称即由此而来。指、趾间有微蹼,具钝爪。背、腹甲的外缘和缘盾腹面均呈米黄色。栖于水域附近,是昼伏夜出的陆生龟类,主要分布于长江以南地区。
黄喉水龟(Mauremys multica)头小,顶部光滑无鳞,吻端超出下颌。腹甲平坦,后缘有一三角形深凹。指、趾间全蹼。尾尖细。咽部及喙黄色,头侧自眼后沿鼓膜上、下各有一条黄色纵纹。栖于我国南方丘陵地带半山区的水域中,常在附近的小灌木及草丛内活动。因其背甲富有胶质,所以常有刚毛藻等附着与之共生,成为“绿毛龟”,民间捕此供作观赏。
(三)海龟科(Chelonidae)背甲扁平,略呈心脏形;肋骨长,末端游离与缘板相连;腹甲各板均小。背、腹甲之间藉韧带相连。头、颈和四肢不能缩入壳内。四肢桨状,指、趾骨扁平而长,有1~2爪。生活于暖水性海洋,以鱼、虾、头足纲、甲壳动物和海藻为食。繁殖季节上岸交配和产卵,每次产卵百余枚;卵壳革质;孵化期40~60天。本科共有6种,我国产4种,常见的是:
海龟(Chelonia mydas)体长约1m,重约300kg~400kg。背甲盾片平整。上颌前端不成钩曲;下颌边缘有锯齿状缺刻。前肢长,内侧有一爪。尾短。繁殖期内的海龟,每年定期成群地游迁到遥远的海边或海岛附近交配。雌龟于夜间爬上岸边沙滩掘坑产卵,产毕以沙覆盖后即回归海洋,龟卵借阳光的热量进行孵化。每年产卵多次,一次产卵90~160枚。生活在太平洋的海龟于夏秋两季常来我国西沙群岛进行繁殖产卵。
玳瑁(Eremochelys imbricata)吻侧扁,上颌钩曲似鹰嘴。幼龟背甲排列成覆瓦状,随同年龄增长渐趋平铺状。前肢有2爪。以海洋动物和海藻为食。背甲艳丽带有光泽,可加热熔化,加工成各种工艺品;分布在我国南海和东海。
(四)棱皮龟科(Dermochelyidae)背甲最大长度可超过2m,重800kg,是现存的最大龟类。背甲由几百枚多边形的小骨板镶嵌而成,外覆革质皮肤,无角质盾片。背甲上有7条纵棱,全部纵棱在体后汇合成一个尖形的末端。四肢桨状,无爪。仅1属1种,即棱皮龟(Dermochelys cori-acea),产于热带和亚热带海洋,我国东海及南海都有分布。
(五)鳖科(Trionychidae)中、小型淡水龟类。背、腹甲骨质,无缘板和角质盾片,覆有柔软的革质皮肤;背甲边缘为厚实的结缔组织,俗称裙边。吻端尖出成可动的吻突,鼻孔开口于此。四肢不能缩入壳内,指、趾间的蹼大,内侧3指、趾有爪。本科约有20多种,主要分布在非洲、东南亚和北美。我国有4~5种。
鳖(Pelodiscus sinensis)又名甲鱼或团鱼。吻突长度约与眼径相等。体较小,最大重量7kg~8kg。咽部有突起,满布血管,是适于在水中交换气体的辅助呼吸器官,因此鳖能在水底潜伏长达10小时之间,不必到水面呼吸空气。除西北地区外,分布遍及全国各省。
斑鼋(Pelochelys maculatus)大型鳖类,体重可达109kg。吻突短,吻长小于眼径的一半。生活在江河和湖泊中,杂食。产于长江下游的江苏南部和浙江北部和云南的东南部,为濒临绝灭的珍稀动物。
三、蜥蜴目(Lacertiformes)
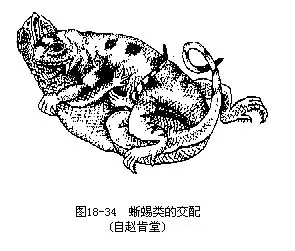
蜥蜴目是爬行纲动物中种类最多的一个类群,多数种类四肢发达,指、趾5枚,末端有爪,适于爬行和挖掘;少数种类四肢退化或缺失。有肩带及胸骨。眼睑可动。舌扁平形,能伸缩,但无舌鞘。左、右下颌骨以骨缝连接,两颌附生端生齿或侧生齿。鼓膜、鼓室及耳咽管一般均存在。陆栖,也有树栖、半水栖或穴居种类。繁殖季节,雄蜥常以袭击方式追逐异性,强行交配(图18-34)。除南极洲外,广布于全球。现存的蜥蜴目动物约3750种,分隶于16科,我国已知约有150多种。代表性种类有(图18-35):

(一)壁虎科(Gekkonidae)皮肤柔软,体被粒鳞。眼大,瞳孔常垂直,无活动性眼睑。有攀援能力的种类,指、趾末端具膨大的吸盘状(指)趾垫,垫的表面有1~2列横形排列的趾下瓣,其上附生着繁多的垂直形细丝,用以扩大与攀援物的接触面。细丝顶端有分泌物,可增加与攀援体的附着力,有利于在平滑的岩石和墙壁上爬行。椎体双凹型。尾有自残现象,再生力强。生活环境多样,夜间活动,能发出叫声。本科约600余种,分布在各大洲的热带及温带地区,我国产30种,代表种类有:
多疣壁虎(Gakko japonica)俗名壁虎和守宫,是长江以南地区习见的一种壁虎,第一指、趾无爪,指、趾下瓣单行;指、趾间无蹼。体背的疣鳞小而密集。与多疣壁虎近似的种类还有分布于华北地区的无蹼壁虎(Gekko swinhonis),它的特征是背面有少量的大疣鳞。
大壁虎(Gekko gekko)体长20cm以上。头及体的背面被粒鳞,并杂有排列成行的大疣鳞。大壁虎干制入药,是传统的著名中药材,因生活时能发出“蛤—蚧”鸣声而得名蛤蚧。分布在广西、广东、福建、台湾和云南等省。
西域沙虎(Teratoscincus przewalskii)头、眼均大,颊部突出。头背为粒鳞,身上和四肢被蜂窝形小鳞,尾上有覆瓦状大鳞。指、趾末端具爪,不膨大成吸盘状,两侧鳞片尖出形成栉状缘。体黄白色,背部和尾上饰有鲜艳的棕黑色横纹。栖于荒漠沙地或戈壁滩,分布在内蒙古、甘肃和新疆,是西北地区的药用蜥蜴。
(二)鬣蜥科(Agamidae)中小型蜥蜴。头背无对称大鳞,体鳞覆瓦状排列,常带棱或有鬣鳞。端生齿,有异形分化趋势。椎体前凹型。尾长,无自残能力。卵生或卵胎生。本科约有300种,主要分布于亚洲。我国产40多种,常见种类有:
斑飞蜥(Draco maculatus)体稍侧扁。体侧有翼状皮膜,为5条延长的肋骨所支持,能滑翔于树枝间,捕食昆虫。咽喉下方具黄色喉囊。行动敏捷,善斗,常成对在一起活动;地上产卵2~5枚。分布于广西、广东、云南和海南岛。
喜山鬣蜥(Agama himalayana)体长约30cm。枕部、颈部和鼓膜下方有锥鳞;背部被弱棱鳞,覆瓦状排列,腹鳞平滑,横列。背面灰棕色,自首至尾有灰白色横纹及纵列圆斑。分布于西藏和新疆,生活在海拔2300m~4100m的山间岩缝及乱石间,也可出现在海拔1400m的荒漠草原上。常于悬崖山腰上挖洞而居,在洞外受惊则迅速窜回洞内或近畔石缝中。有时也在树下穴居,白天爬上树去,捕吃昆虫。
草原沙蜥(Rhrynocephalus frontalis)头形似蟾,上、下睑鳞的边缘锯齿形,闭眼时可阻止沙石伤及眼睛。无鬣鳞。指、趾两侧有发达的栉状缘,可增大四肢与地面接触的面积,免于蜥体陷身沙内。有将尾甩到背部作上下卷动的习性。遇惊时,常疾驰而逃,或在沙地里急速摇摆身子,使蜥体下沉,同时用后肢交替挖掘细沙,容扬起之沙落于体背而隐身其中。卵生,7月孵出幼蜥,分布在内蒙古、河北、陕西、甘肃、宁夏和北京地区。
(三)石龙子科(Scincidae)头顶有对称排列的大鳞片。全身被覆瓦状圆鳞,角质鳞下具有真皮性骨板。侧生齿,尖出或钩状。尾粗圆,有自残能力。卵生或卵胎生。本科约有600余种,广泛分布在各大洲。我国产31种,代表性种类有:
蓝尾石龙子(Eumeces elegans)体形纤细,四肢较短。体鳞圆而光滑,背面深黑色,有 5条黄色纵纹,直达尾部,尾的后半部蓝色。栖于低山山林及山间道旁的石块下,亚卵胎生繁殖。产于我国长江以南各省。
蝘蜓(Lygosoma indicum)体长约90mm,短于尾长。因体呈古铜色而俗称铜石龙子,体侧各有一条醒目的黑色纵纹,止于尾基。生活于平原或低山地区,卵胎生,每次产仔6~8尾。分布在河南、陕西、甘肃和长江以南各省。
(四)蜥蜴科(Lacertidae)头顶有对称的大鳞片,腹鳞方形或矩形,纵横排列成行。四肢发达,有股窝或鼠蹊窝。尾长易断,也易再生。生活于林区草丛、草原、荒漠和平原地带。卵生或卵胎生。全世界约有140余种,广泛分布在欧、亚、非三洲,我国产20种,代表性种类有:
北草蜥(Takydromus septentrionalis)身体和尾均甚细长,头长吻尖,有颅顶孔,耳孔明显,鼓膜外露。四肢和指、趾也长,大腿内侧基部各一个鼠蹊窝。背部及腹面被方形棱鳞,平行排列。体棕绿色或鲜绿色,分布于我国东部和西南各省。
丽斑麻蜥(Eremias argus)体较短粗,四肢短小,尾长与头体长大致相等。背鳞颗粒状;腹鳞方形,无棱,向腹中线排成斜行。咽部有一横褶形的领围。大腿内侧各有一列股腺窝。背部及腰侧有纵列的白色眼斑或链形长条。常在长有灌木丛或芨芨草堆周围的沙地上捕食昆虫。分布在我国东北、华北和西北(不包括新疆和青海)各省。
(五)蛇蜥科(Anguidae)体呈蛇形,有后肢骨的残余。全身被覆瓦状圆鳞,鳞下有真皮性骨板。眼小,有活动眼睑。体侧有凹入的纵沟。尾长易断,能再生。侧生齿,形状不一。本科有50种左右,我国产3种,最常见的是脆蛇蜥(Ophisaurus harti),生活在草丛中,穴居,夜间外出活动,以昆虫和蜗牛等软体动物为食。行动方式似蛇,但甚缓慢,雌性有盘匍在卵上的习性。分布在长江以南各省。
(六)鳄蜥科(Shinisuridae)体形似鳄,躯干粗壮,四肢发达,指、趾末端具弯曲的利爪。尾长而侧扁。头顶鳞片形状不一,大致对称。背部的粒鳞间杂有大形棱鳞,并形成纵列往后延伸至尾部呈2行棱嵴。鼓膜不显。舌短,前端分叉。侧生齿。卵胎生。本科仅一种,即产于我国广西瑶山的鳄蜥(Shinisaurus crocodilurus),是极其珍贵的爬行动物,为国家一级重点保护动物。
(七)巨蜥科(Varanidae)体型巨大,是蜥蜴目个体最大的类群。身体及四肢均甚粗壮,尾长而侧扁。头顶无对称的大鳞片,背面被粒状棱鳞,腹鳞方形,鳞下有真皮性骨板。舌细长,分叉,可缩入舌基的鞘内。侧生齿。尾无自残能力。本科仅1属30种左右,分布于非洲、大洋洲、亚洲南部及东南部。我国产圆鼻巨蜥(Varanus salvator),一般都在水域附近生活,陆栖,能在水中游泳,也可攀援上树觅食。肉食性。夏天雨季开始产卵,卵产在岸边的土穴内或树洞中,约15~30枚。分布于云南、广东、广西和海南省。
(八)避役科(Chamaeleontidae)形似蜥蜴,但因适于树栖生活而各种器官的构造极端特化。本科共3属,85种,除了有2种分布在西班牙南部、印度和斯里兰卡外,其他种类全产于非洲和马达加斯加岛。本科的代表是避役(Chamaleon sp.),身体侧扁,被覆粒鳞,背部有脊棱。四肢长,前、后脚的5枚指、趾分成互相对生的两组:内侧3指愈合成一组,外侧两指愈合成另一组;第一、二趾形成一组,其他3趾形成一组,适于抓握树枝。尾长,善于缠绕。眼大而凸出,左、右两眼都能独立地往各个方向转动,搜索捕食目标,视野广阔,发现昆虫后可作进一步的对光调节,直到双眼同时瞄准虫体。舌长,末端膨大,富有粘液,捕虫时能迅速射出,投粘猎物。真皮层有色素细胞、鸟嘌呤结晶和折光性极强的微粒,在神经支配下,能快速改变体色适应周围环境,“变色龙”的俗名即由此而来。卵生或卵胎生(南非的侏儒避役)。
(九)毒蜥科(Helodermatidae)唯一有毒的蜥蜴,牙齿弯曲而基部膨大,下颌齿的前、后两面均有深沟,下唇腺特化成毒腺。体形肥胖,尾短而粗,背面有珠状小瘤,皮下有扁平的骨鳞。体色醒目可怖,背部灰白色或黑色,饰有斑驳错落的粉红色、黑色及黄色的斑点,尾具宽阔的深色环纹。栖于沙地,行动迟缓,以蚁类、蛇卵及小鼠为食。卵生。本科仅1属2种,即短尾毒蜥(Helo-derma suspectum)和珠背毒蜥(Heloderma horridum)。
四、蛇目(Serpentiformes)
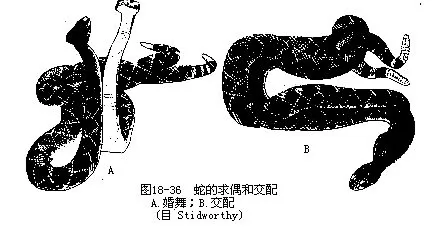
体细长,分头、躯干和尾三部,颈部不显。四肢消失,带骨及胸骨退化,但蟒科、盲蛇科、瘦盲蛇科和筒蛇科(Aniliidae)的种类还有骨盆及残余后肢。无活动性眼睑、瞬膜和泪腺。缺乏鼓膜,鼓室萎缩,耳咽管也消失;内耳的卵圆窗和方骨之间由耳柱骨相接。颅骨由于大量膜骨退化或消失,已无双颞孔的痕迹。椎体前凹型。除寰椎和尾椎外,其余椎骨上都附有可动的肋骨,是蛇类向前爬动的主要支持器官。成对的内脏器官因受体形的影响,使其左右对称的位置变换成前后交错的位置,或只保留一侧器官,另一侧则萎缩、退化。无膀胱。雄性有一对交配器。卵生或卵胎生。交配前有求偶的婚舞行为(图18-36)。
蛇类是蜥蜴在进化过程中高度特化的一个分支,在种系发生上与蜥蜴的亲缘关系比较密切,所以蛇的有些特征也同样见于蜥蜴的某些种类中,例如蛇蜥科没有四肢,体呈蛇型;巨蜥科的舌尖分叉和蛇一样;鬣蜥科的耳孔不显与蛇相似等。全世界现存蛇类约有3200种,分隶于13科,广布于各大洲。我国产210多种,其中有毒蛇50种。代表种类有如下科(图18-37):
(一)盲蛇科(Typhlopidae)包括一些形似蚯蚓的小型无毒蛇类。周身覆有形状相同的圆鳞,光滑无棱。眼甚退化,隐于鳞片之下,故称盲蛇。因方骨不能活动和下颌骨左右两半在前端愈合,所以无法扩大张口角度。上颌有少量牙齿,下颌无齿。有腰带骨的残迹,表明本科属于较原始的蛇类。卵生或卵胎生。我国产3种,最常见的是钩盲蛇(Ramphotyphlops braminus),长约15cm,是我国蛇类中最小的一种。营穴居生活,吃蚯蚓、白蚁、蚁卵、昆虫及其幼虫。分布在长江以南各省。
(二)蟒蛇科(Boidae)背鳞小而光滑,腹鳞大而宽阔。腰带骨退化,但仍留有股骨残余。泄殖孔两侧有一对角质爪状物,有成对的肺。卵生或卵胎生(沙蟒)。有的卵生种类具有孵卵行为,母蟒借肌肉节律性收缩能升高体温,有助于卵的孵化。主要以恒温动物为食,大多数种类发展了与这种食性相适应的热能感受器。本科约有60余种,分布于热带和温带地区。我国南方的蟒蛇(Python molurus)和西北的沙蟒(Eryx miliaris)为本科的代表。
(三)游蛇科(Colubridae)本科是蛇目中包括种类最多的类群,现存蛇类种数的2/3都属于这一科。头顶有对称大鳞片,腹鳞宽大。两颌都有牙齿,少数种类的上颌骨后端有2~4枚较大的沟牙。卵生或卵胎生。分布几遍全球,我国产140多种,常见的有:
黄脊游蛇(Coluber spinalis)全长约0.5m。背面褐绿色,自额部沿脊梁至尾端有一条明显的浅黄色纵带。是黄河以北的优势种蛇类之一,往南可达长江下游各地,大多以蜥蜴和鼠类为食。
赤链蛇(Dinodom rufozonatum)是常见的无毒蛇。背面黑色,有70余条红色横纹;头部鳞片黑色而鳞缘绯红,颅顶至项背有一人字形红纹;腹鳞浅黄无斑。以鱼、蛙、蟾蜍、蜥蜴、幼鸟和小鼠为食。分布于华北、东北和长江以南各地。
黑眉锦蛇(Elaphe taeniurus)全长达1.5m以上。眼后有一黑纹延向颈部,状如黑眉而得名。常栖于农舍及其附近的园地、树洞,性较凶暴,受惊时即竖起头颈准备攻击,大量捕食鼠类,是著名的益蛇。分布于东北、华北和长江以南各省。
虎斑游蛇(Rhabdophis tigrina)背面暗绿色,身体前段两侧有交互相间的黑色和桔红色斑块,往后则红色逐渐消退;腹面青灰色。生活在水边草丛,捕食蛙、蟾和鼠类。
乌梢蛇(Zoacys dhumnades)又名乌蛇和乌风蛇。最大的个体全长可达2m以上。背部绿褐色或黑褐色,中央2~4行背鳞起棱,两侧有2条纵贯全身的黑纹,但成年个体在后部逐渐变得隐晦而不显。生活于丘陵地带及田野间,主要以蛙为食,也吃鱼和蜥蜴。分布于长江以南地区,是我国的传统中药材。
中国水蛇(Enhydris chinensis)又名泥蛇。蛇体圆柱形,头形略扁而宽于颈部;背鳞平滑;尾短,尾下鳞双行。鼻孔朝上,适于半水栖生活。最后2枚上颌齿为较大的沟牙。背面褐绿色,外侧第二、三行鳞片白色,形成纹条。栖于稻田、池塘、水沟等处,捕食鱼类。分布在长江以南地区,是本科中的有毒蛇类。

(四)眼镜蛇科(Elapidae)外形上与一般无毒蛇不易区别,上颌骨的前部有一对较大的前沟牙,其后尚有几枚预备毒牙。本科的蛇毒主要作用于人和动物的神经系统,称神经毒类。全世界的毒蛇中,有一半左右的种类都隶属于本科,约180多种,分布在大洋洲、亚洲、非洲和美洲。我国有9种,全分布于长江以南地区。常见种类有:
眼镜蛇(Naja naja)全长1m有余,背部黑褐色,有狭窄的黄白色横斑。惊怒时,蛇体前部能昂竖直立,呼呼作声,频频向前摆动作攻击状,并膨大颈部,使该处的白色斑纹呈现眼镜状,故名。栖于平原及山地森林中,夜间活动,捕食鱼、两栖动物和雏鸟等。分布在华南各省,是凶猛的毒蛇之一,也是引起蛇伤的主要蛇种。与眼镜蛇近似的还有眼镜王蛇(Ophiophagus hannah),全长2m~6m,主要特征是有一对大枕鳞。
银环蛇(Bungarus multicinctus)全长1m左右。头部椭圆形,略大于颈;尾梢尖细。体表具黑色和白色相间的环纹。既是引起蛇伤的主要蛇种之一,又是浸泡“三蛇酒”和“五蛇酒”等药酒的原料之一,干幼蛇称金钱白花蛇,为著名的中药材。分布于我国长江以南各省。近似种有金环蛇(Bungarus fasciatus),体表有黑色和黄色相间的环纹。
(五)海蛇科(Hydrophiidae)本科全是终生生活于海洋中的前沟牙类毒蛇。体后部及尾侧扁;鼻孔位于吻背,有启闭自如的鼻瓣;腹鳞窄或消失;卵胎生;以鱼类为食。本科共49种,分布于印度洋和太平洋温水区域。我国沿海产16种,分布最广是长吻海蛇(Pelamis platurus),蛇体背黑腹黄,截然分界,故别名黄腹海蛇。尾部可有5~10块黑斑,尾端黑色。海栖,吃小鱼和甲壳类动物。分布在我国的黄海、东海和南海沿海。
(六)蝰科(Viperidae)上颌骨短而高,附生着长而弯曲的管牙及若干副牙。由于头骨的机械活动,闭口时上颌骨连同管牙卧于口腔顶部;张嘴时上颌骨和管牙都一起竖立起来。全是毒蛇,蛇毒为血循毒类,主要作用于心血管系统及血液。根据头部有无颊窝和顶鳞的大小,可分为蝮亚科(Crotalinae)、蝰亚科(Viperinae)和白头蝰亚科(Azemiopinae)。本科共有180余种,在地理分布方面,蝮亚科分布在美洲、亚洲和欧洲,但无蝰亚科种类,而蝰亚科的分布中心在非洲(欧、亚两洲也有),但缺乏蝮亚科蛇类。白头蝰亚科仅1属1种,即白头蝰(Azemiops feae),仅见于我国和越南。我国常见的蝰科蛇类有:
蝮蛇(Agkistrodon halys)头略呈三角形,颈部明显。有颊窝。背鳞具棱。背部颜色变化于灰褐色到土红色之间,交互排列着黑褐色圆斑或波状横斑。眼后有黑色条斑,其上方镶有黄白色细纹。主要以脊椎动物为食。卵胎生,每次产仔蛇2~15条。分布几乎遍及全国。
尖吻蝮(五步蛇)(Deinagkistrodon acutus)体长达1.5m。吻端尖出并向上方翘起,又名蕲蛇;颊窝明显。背面灰褐色,布有灰白色的菱形方斑,腹面白色,有黑斑,俗称白花蛇和棋格蛇。卵生。主要分布在长江以南地区。
烙铁头(Trimeresurus jerdonii)全长1m左右。颊窝显著。背面草绿色,杂有黄、红及黑色斑点,红斑在脊背正中形成一行较大的斑块。栖于海拔1700m~3100m的山区石堆或杂草中,是川西牧区的剧毒蛇,吃鸟和小型兽类。卵胎生。近似种有竹叶青(Trimeresurus stejnegeri),头呈三角形,颈细。周身绿色,体侧自颈至尾有一条鲜明的黄线或白线,易于辩识,是造成蛇伤的剧毒蛇之一。两者都分布在黄河以南地区。
草原蝰(Vipera ursini)无颊窝。头部背面鳞片光滑。背部灰褐色,背脊正中有一行黑褐色锯齿状纵纹,体侧有暗褐色斑点缀成的纵纹2~3行。生活在草原、疏林和芦苇丛,也见于海拔3000m的山区。吃蝗虫(占食物总量的90%以上)和蜥蜴。卵胎生,产1~6条仔蛇,最多可产17条。分布于新疆,是当地的主要毒蛇。
五、鳄目(Crocodiliformes)
鳄目心脏有2个完全隔开的心室;头骨有发达的次生腭;两颌有槽生齿;具横隔。颅骨双颞孔型。腹壁内有游离的腹膜肋。
现存鳄目动物共22种,分别隶属于鳄科(Crocodilidae)、鼍科(Alligatoridae)和食鱼鳄科(Ga-vialidae),它们大多生活于非洲、大洋洲、亚洲南部及热带美洲等温暖地区。我国长江中、下游的扬子鳄及美洲的密河鳄是鳄类过去在北方温带分布的唯一孑遗。
扬子鳄(Alligator sinensis)是小型鳄类之一,全长约2m,中国古藉中称为鼍。吻短而钝,下颌第四齿大,闭嘴时嵌合于上颌的凹槽内。皮肤革质,覆有角质方形大鳞。前肢5指,后肢4趾,内侧3指、趾具爪。泄殖腔孔纵裂。栖于江湖岸边的滩地,芦苇或竹林丛生处。雌雄分别挖穴而居,洞内通道曲折起伏,可长达数10m,呈复杂的迷宫状。11月至次年4月为冬眠期,4~5月苏醒出洞。6~7月进行交配和筑巢产卵,卵数约20枚,最多时可达52枚。亲鳄有护卵习性,卵的孵化期为60~70天,夜间觅食,食物中的主要成分是田螺和螺蛳,占到总食量的63%,也吃鱼、蛙、虾、蚌和小兽等。
扬子鳄是我国的特产,目前它的分布区仅限于安徽省长江以南、皖南山系以北的丘陵地带,以及江苏省和与皖南交界的浙江省一角,全部野生个体不足500尾。为我国一级重点保护动物。1982年,我国在安徽省南部的宣城成立了扬子鳄繁殖研究中心,至今人工繁育的扬子鳄幼鳄数已达到数千尾之多,为拯救国宝和使世界濒危动物摆脱绝灭的命运,作出了巨大贡献。
马来鳄(Tomistoma schlegelii)是分布在马来西尼和印度尼西亚的大型淡水鳄类。它在一千多年前曾广泛地生活于我国广东、广西两省,并且由于数量众多、性猛凶恶、袭食人畜而是当地的一害。唐代广东潮州(今潮安)刺史韩愈曾写下名作《祭鳄鱼文》,文中所指的即马来鳄。该鳄在气候变化、人口密集、开垦务农和破坏栖息生境的情况下,又遭到人们的恣意杀戮,终于在元、明两代期间绝灭殆尽。有人认为我国产湾鳄(Crocodilus porosus),且古藉中也有在海南岛至西沙群岛一带见过咸水鳄的记载,所以南海沿海过去有过少量湾鳄,也是有可能的(图18—38)。
第三节 爬行动物与人类的关系
一、爬行动物的益处
(一)生态系统中的作用爬行动物为外温动物,主要依靠吸收太阳的辐射热来维持和提高体温。由于新陈代谢率低,对自然界内作为热量来源的营养物质消耗也少,它们所摄入的大部分能量都能通过同化作用而转变为自身的生物量,其净生产力可达到30%~90%,远远超过恒温动物。
大多数爬行动物都是杂食或肉食类,蜥蜴和蛇类通过大量捕食昆虫及鼠类等摄入能量而有益于农牧业生产,在生态系统中充当着次级消费者的角色。据对有益系数的调查,蓝尾石龙子为53.69%,石龙子为51.6%,蝘蜒为24.01%,而分布极广的草原沙蜥和密点麻蜥,分别达到82.14%及81.10%。许多爬行动物又是食肉兽和猛禽的食物及能量的来源之一,在生态系统能量的流转过程中,又处于次级生产力的地位。因此,爬行动物对维持陆地生态系统的稳定性,以及为自然界提供能量贮存来说,具有不可忽视的作用。
(二)食用蛇肉味美,富有营养,含脂肪22.1%,蛋白质18%,并有多种氨基酸成分,所以是对身体有滋补和治病作用的食品。两广地区以眼镜蛇、金环蛇和灰鼠蛇为原料制作生产的三蛇菜、三蛇酒、三蛇胆都是名闻遐迩的著名酒食。鳖甲周缘的裙边,历来就是脍炙人口的佳肴,并为名贵的滋补食品。海龟肉、龟蛋、鳍脚、脊肌、腹甲骨片缝间的黄脂肪等是太平洋上许多岛屿居民的喜爱美食。目前由于乱捕滥猎,已导致资源严重破坏,必须加以控制。
(三)工艺用蛇皮的皮质轻薄,富有韧性,花纹美观,不但可以制作皮革、皮带、皮鞋、提包、钱袋等工艺品,也能用作胡琴、手鼓、三弦的琴膜及鼓皮等民族乐器。
(四)医药用我国用于入药的蜥蜴至少有10多种,其中最负盛名的是产于广西的大壁虎,药名为蛤蚧,据《本草纲目》记载有补肺气、益精血、定喘止咳、疗肺痈消渴、助阳道的功能。以蜥蜴作为蛤蚧代用品的在西藏和新疆有鬣蜥,内蒙古有荒漠麻蜥,甘肃省有西域沙虎等。鳖甲和龟板、蛇肉、蛇胆、蛇蜕、蛇毒都可入药。蛇胆可加工成蛇胆川贝液、蛇胆陈皮末、蛇胆半夏液等中成药,治风湿关节痛、咳嗽多痰等病。蛇蜕的中药名叫龙衣,入药有杀虫祛风的功能,可治疗喉痹疔肿、疥癣和难产。
蛇毒研究是生物科学中极有开发前途的一个分支,我国对蛇毒研究已由蛇毒血清的试制,逐步深入到有关蛇毒的生化及其综合利用方面。目前已制成的眼镜蛇毒注射剂具有比吗啡更有效和更持久的镇痛作用,对于减轻晚期转移癌痛、三叉神经和坐骨神经痛、风湿性关节痛、脊髓痨危象、带状疱疹等病人的剧痛,都有明显的效果。用蛇毒酶治疗癌症也收到了一定疗效。蝰蛇蛇毒有较强的凝血性,对于机体缺乏凝血的血友病患者,可用蛇毒治疗其出血性疾病的局部止血。我国学者从蝮蛇蛇毒中提取的抗栓酶,在临床已用于脑血栓、血栓闭塞性脉管炎、冠心病的治疗。
(五)其他方面 蛇类对地壳内部的剧烈震动、地温升高及地面发生反复无常的倾斜运动等,具有很强的敏感性,因而可能在地震前表现出反常的行为。随着仿生学的发展,科学家还根据毒蛇颊窝的构造及其独特的热测位器作用,把研究成果应用到红外线测位仪上,并制成具有高度精确性和能追踪飞机、潜艇、车辆的响尾蛇导弹及火箭自导装置等。从海龟洄游路线的导航机制可启发改善航海仪器的研究。也有人认为龟类背甲符合最优结构的薄壳结构理论,在大型建筑设计上有借鉴之处。
我国的爬行动物资源丰富,有许多举世闻名的扬子鳄和鳄蜥等特产种类,对于科学研究和国际文化交流都有重要意义。
二、毒蛇的危害及蛇伤防治
(一)毒蛇与无毒蛇的区别在现存的蛇类中,约有近600种为毒蛇,我国产毒蛇50种左右。
蛇毒是一种复杂的蛋白质,进入人和动物体内后,能随淋巴及血液扩散,引起中毒症状。前沟牙蛇类的蛇毒对人的危害较大,分别含有眼镜蛇神经毒、α-环蛇毒神经毒和海蛇神经毒等神经毒(neurotoxic),可引起乙酰胆碱失去作用,造成机体的神经肌节头之间的冲动传导受阻,短时间内导致中枢神经系统麻痹而死。管牙类的蛇毒中含有血循毒(hemotozic),可引起伤口剧痛、水肿、渐至皮下出现紫斑,最后导致心脏衰竭死亡。毒蛇毒液的主要成分见表18—1。

通常蛇毒的毒性强度与各种蛇毒的性质,以及毒蛇咬人时的排毒量有关。例如,眼镜王蛇排毒量多,毒性强;竹叶青排毒量少,毒性也强;银环蛇排毒量少,但毒性强,了解这些对于有效地预防被毒蛇咬伤和处理蛇伤,具有重要的积极意义。
毒蛇与无毒蛇最主要的区别,在于毒蛇有毒腺和毒牙,无毒蛇则无此特征。此外,还有下列各点可供识辩毒蛇和无毒蛇(表18—2,图18—39,图18—40):

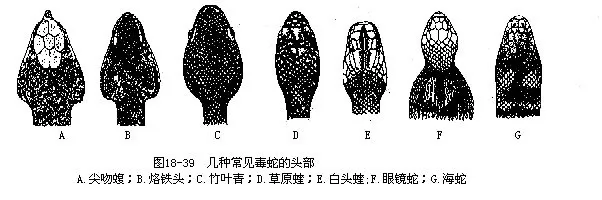
我国的毒蛇中,有10种属于游蛇科的后沟牙类,并不对人造成危害。海蛇科的毒蛇约有16种,终生生活于海域中,只有沿海的渔民偶而被咬的病例,其他20种毒蛇中,至少又有13种十分罕见或分布区极其狭窄。所以毒性较强、分布较广、数量较多和经常引起蛇伤的毒蛇,通常只占毒蛇种类的1/5左右。长江以北广大地区内的主要毒蛇是蝮蛇。包括四川、云贵高原及横断山脉的西南地区,主要毒蛇有蝮蛇、尖吻蝮和几种烙铁头。由长江中、下游沿岸,向南到达南岭山脉的华中地区,丘陵地带有蝮蛇、眼镜蛇及银环蛇,而山区则有尖吻蝮和竹叶青等。南岭山脉及其以南的华南地区,是我国蛇种甚多的区域,主要毒蛇有眼镜蛇、银环蛇、金环蛇、眼镜王蛇、尖吻蝮、烙铁头和竹叶青等。青藏高原的主要毒蛇有眼镜王蛇、高原蝮、墨脱竹叶青、西藏竹叶青及菜花烙铁头等。
(二)蛇伤及其防治 毒蛇咬伤最严重的地区是南美、南亚和东南亚各国。据报道,世界上每年约有数十万人被毒蛇咬伤。印度在1968年的蛇伤总人数高达30万~40万人,死亡的就有3万多人。同样,在南亚次大陆和我国也有相当数量的蛇伤病人,可见毒蛇咬伤在这些地区的危害是非常严重的。此外,草原牧场(特别是早春季节)的毒蛇常对畜群造成伤害,例如我国新疆伊犁地区的蝮蛇、草原蝰和阿勒泰地区的极北蝰等毒蛇,都危害人畜安全。过去因蛇害致死的马匹,约占总死亡数的20%,数量相当可观。近年来我国动物学工作者与当地群众相结合,采用生态学和药熏方法防治蛇害,取得显著成绩。

一般来说,毒蛇的行动大多比较迟缓,很少主动攻击咬人,只有当人们在砍柴、割草、秧、夜行中,无意踩到或接触蛇体时才会发生咬伤事故。因此,蛇伤的部位通常都在下肢的脚踝以下,其次是上肢或头、胸部。蛇类活动的最适气温是18℃~30℃,所以在我国长江以南地区。7~9月是蛇伤发病率最高的季节,尤其在夏季闷热欲雨或雨后乍睛的天气,由于蛇洞内气压低而湿度大,毒蛇经常出洞活动,咬人致伤。
如果被毒蛇咬伤,在条件许可下应立即将蛇击毙,同时将蛇带往就医,这对根据毒蛇的种类来采取对症治疗是极为必要的。假如确系毒蛇所咬,就会在伤处留有2个大而深的牙痕,发红的伤口灼热疼痛,在几分钟内显著地肿胀起来,并迅速扩展肿胀范围,同时还会发生头晕、眼花、抽搐、昏睡等症状。
毒蛇咬伤的紧急局部处理原则是尽快排除毒液,延缓蛇毒的扩散,以减轻中毒症状。一般应立即在伤口上方2cm~10cm处用布带扎紧,阻断淋巴和静脉血的回流,并每隔15~20分钟放松布带1~2分钟,以免血液循环受阻,造成局部组织坏死,如注射抗蛇毒血清后,可解除结扎。结扎后,应用清水、盐水或0.5%浓度的高锰酸钾溶液反复冲洗伤口。此外,还可使用扩创排毒(被尖吻蝮或蝰蛇咬伤不宜采用此法)、拔火罐或口吸法等排除蛇毒。紧急处理后,要及时就近求医治疗。
我国在蛇毒分析和蛇伤防治方面的研究均已取得重大成就。目前,除运用单价及多价抗蛇毒血清和α-糜蛋白酶等特效药物治疗蛇伤外,还采用多种草药及研制成各种蛇药,极大地提高了毒蛇咬伤的治愈率。
复习题
1.简述羊膜卵的主要特征及其在动物演化史上的意义。
2.归纳爬行类适应于陆生的主要特征。
3.简述现存爬行类4个目及主要科的特征
4.总结爬行动物与人类的关系。
5.毒蛇与无毒蛇的区别以及毒蛇防治原则
本文标题:第十八章 爬行纲(Reptile)
手机页面:http://m.dljs.net/dlsk/dongwuxue/59415.html
本文地址:http://www.dljs.net/dlsk/dongwuxue/59415.html
