一、线虫动物门的主要特征
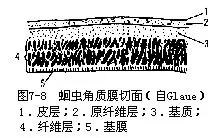
(一)角质膜(Cuticle)线虫体表被一层角质膜,厚度一般为身体半径的0.07倍,坚韧富弹性,主要成分为蛋白质。角质膜为上皮分泌形成,一般分为皮层(cortex)、中层(median)(基质)和基层(basal layer)(斜行纤维)3层①70000034_0150_0,最内为基膜(图7-1),角质膜有保护作用。线虫在生长发育过程中,有几次脱去旧的角质膜,长出新的角质膜,称为蜕皮(ecdysis)。线虫在两次蜕皮间及最后一次蜕皮后均生长。

(二)原体腔(primary coelom)线虫的体壁由角质膜、上皮和纵肌层组成,又称皮肌囊。角质膜下为合胞体(syncytial)的上皮(epidermis),即上皮细胞的界限不清,具多核。上皮向内突起成脊,于身体左右两侧形成侧线(lateral cord),于背侧和腹侧形成背线(dorsal cord)和腹线(ven-tral cord)。上皮内为中胚层形成的纵肌层,不发达,属典型的斜纹肌。
线虫体壁围成的广阔空腔称为原体腔,又称假体腔(pseudocoel),是由胚胎时期的囊胚腔发展形成。原体腔只有体壁中胚层,且不具体腔膜(peritoneum),无脏壁中胚层。原体腔的出现,是动物进化上一个重要特征。线虫的原体腔内充满体腔液,致使虫体鼓胀饱满,内压平均为9.33kPa~13.33kPa(70mm~100mm汞柱),最高可达225mm汞柱。故身体难以任意伸缩,只依靠纵肌收缩,沿背腹向弯曲作波浪蠕动,弯曲波由体前向后传递,而进行游泳或爬行。
(三)发育完善的消化管线虫为发育完善的消化管,即有口有肛门。消化管分为前肠、中肠和后肠三部分(图7-2)。前肠由外胚层于原口处内陷形成,内壁有角质膜,分化为口、口腔及咽。口腔内常形成齿、口针(oral stylet)等,可辅助摄食;咽外壁有发达的辐射状肌肉,收缩时使三角形咽腔迅速扩大,故有吮吸作用。大多数线虫的咽外有单细胞咽腺,能分泌多种消化酶,进行细胞外消化。中肠由内胚层发育形成,为消化与吸收的主要部分。后肠亦为外胚层于胚胎后端处内陷形成,内壁也具角质膜,包括短的直肠和肛门。寄生线虫的消化管简单,有退化趋势,无消化腺。
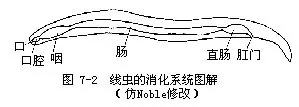
食物由口摄入,在中肠内进行细胞外消化,不能消化的食物残渣,由肛门排出。这样的消化机能更为完善,与胃循环腔相比是个飞跃的进步,这也是动物进化的特征之一。
(四)排泄器官线虫的排泄器官结构特殊,没有纤毛及焰细胞存在,可分为腺型(glandulartype)和管型(tubular type)2种。腺型排泄器官属原始类型,通常由1~2个称为原肾细胞(renette cell)的大的腺细胞构成,海产自由生活种类的线虫(如Linhomeus)属此,但一般为1个原肾细胞,位咽的后端腹面,排泄孔开口于腹侧中线(图7-3A)。小杆线虫(Rhabdias)具有2个肾细胞(图7-3B)。原肾细胞吸收体腔液中的代谢产物排出体外。
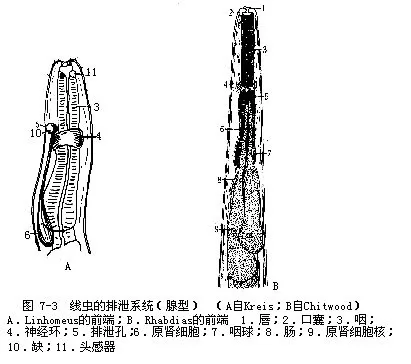
寄生线虫的排泄器官多为管型,是由一个原肾细胞特化形成,由纵贯侧线内的2条纵排泄管构成,二管间尚有一横管(有的呈网状,如蛔虫)相连,略呈“H”型(图7-4)。由横管处伸出一短管,其末端开口即为排泄孔,位于体前端腹侧。溶于体腔液中的代射产物,通过侧线处的上皮进入排泄管。显然管型排泄器官是由腺型排泄器官演变而来。
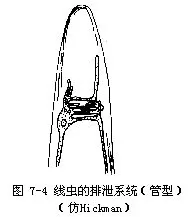
线虫的排泄器官无纤毛和焰细胞,显然不同于扁形动物的原肾管(proto-nephridium),但这种排泄器官也是由外胚层形成,从结构与机能上看,类似原肾系统,可以看成是一种独特的原肾管。
(五)生殖 线虫为雌雄异体(dioecious),且雌雄异形,雄性个体小于雌性个体。有极少数种类为雌雄同体(hermaphroditism),如某些小杆线虫(Rhabditoid)和植物线虫。更有一些种类只有雌虫存在,未发现雄虫,营孤雌生殖(parthenogenesis)。数种陆生线虫和根结线虫虽有雄虫存在,也出现孤雌生殖(如Mermis subnigrescens及Heterodera marioni等)。雌雄同体常称为共殖(syngony),共殖线虫的外形呈雌性,在共殖线虫培养中,偶然也有雄虫出现。
线虫的生殖器官为细长管状,雄性为单个(Heterodera marioni及Anticoma typica成对),分化成精巢、输精管、储精囊、射精管,通入直肠,精子由肛门排出,故直肠实为泄殖腔(colaca),肛门即泄殖孔。雌性的生殖器官成对,有卵巢、输卵管、子宫,2条子宫汇合成一短的阴道,开口于腹侧中线上的雌性生殖孔。
精子的形态因种类不同而异,一般无尾,呈圆形,可作变形运动。卵具壳。雌雄虫交配,卵在子宫内受精,个体发育中有幼虫阶段。生长过程中有蜕皮现象。自由生活的种类产卵量小,寄生线虫产卵量巨大。
(六)神经系统线虫的神经系统有围绕咽部的围咽神经环(circumen-teric ring),主要是神经纤维,只有少数神经节细胞(ganglion cell)。与围咽神经相连的主要神经节有成对的侧神经节(lateral ganglion)和单个或成对的腹神经节(ventral ganglion)。神经环向前后伸出多条神经,均嵌在上皮中,以背神经和腹神经最发达。
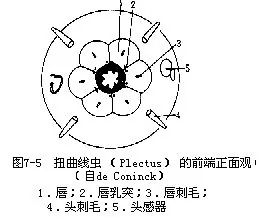
线虫的感官不发达,头端有头刺毛、唇乳突,为触觉器官;头感器(am-phid)可接受化学刺激(图7-5)。尾端有尾乳头、尾感器(phasmid)。寄生线虫的头感器退化,尾感器发达。
二、代表动物——人蛔虫(Ascaris lumbricoides)
人蛔虫是人体最常见的肠道寄生线虫之一,感染率高,尤其是儿童。人蛔虫与猪蛔虫(A.suum)二者形态结构非常相似,染色体均为2n=24;DNA含量也无显著差异(p>0.05)(黄跃进等,1988)。故有时也用猪蛔虫作代表动物。
(一)外形人蛔虫体呈圆柱形,向两端渐细,全体乳白色,侧线明显。雌虫长200mm~250mm,直径5mm左右;雄虫较短且细,尾端呈钩状(图7-6)。虫体前端顶部为口,有3片唇,背唇一片,具二双乳突(double papillae),腹唇2片,各具一双乳突和一侧乳突(lateral papilla)(图7-7)。口稍后处腹中线上有一极小的排泄孔。肛门位体后端腹侧的中线上。雌性生殖孔在体前部约1/3处腹侧的中线上,很小;雄性生殖孔与肛门合并称泄殖孔,自孔中伸出一对交合刺(spicule),能自由伸缩。

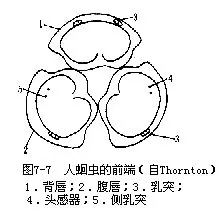
(二)构造
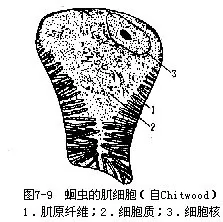
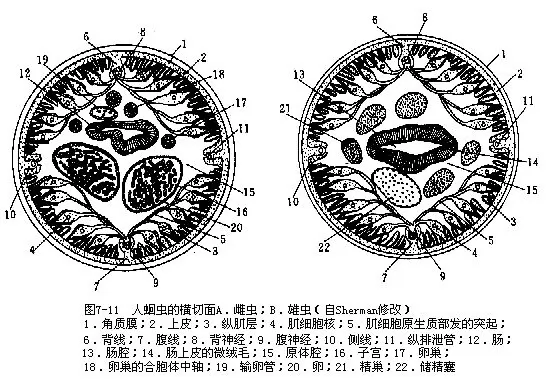
1.体壁及原体腔人蛔虫的体壁由角质膜、上皮和肌层构成皮肌囊。角质膜发达,由皮层和原纤维层(fibril layer)(属于皮层)、基质(matrix)(中层)、纤维层(fiber 1ayer)(基层)及基膜构成,有保护作用(图7-8)。上皮层为合胞体构造,两侧线发达,其内各有一条纵排泄管;背线及腹线明显,内有背神经和腹神经。纵肌不发达,为背线、腹线及侧线分成4条纵带(图7-11),故皮肌囊不完整。肌细胞基部具肌原纤维,端部为原生质部分,细胞核即位此部(图7-9)。
体壁内为广阔的原体腔,内充满体腔液,虫体饱满鼓胀,纵肌伸缩时只能作弯曲的蠕动,消化管及生殖器官浸在体腔液内(图7-11)。
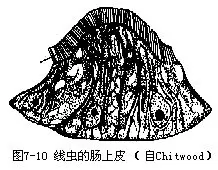
2.消化系统消化管简单,为一直管(图7-14),口腔不发达,口后为一肌肉性的管状咽,内腔呈三角形,外壁的辐射状肌肉发达,有吸吮功能。咽后为肠,肠壁为单层柱状上皮细胞构成,内缘有微绒毛(microvilli)(图7-10)。直肠短,以肛门开口于体外。雄虫的直肠实为泄殖腔,以泄殖孔开口。蛔虫无消化腺,它摄取的食物是宿主肠内已消化或半消化的物质,一般可以直接吸收。
3.呼吸与排泄蛔虫生活在含氧量极低的肠腔内,行泛氧呼吸,即借酶的作用,分解体内储存的糖原,以获得能量。故泛氧呼吸为寄生线虫的特点之一。
蛔虫的排泄器官属管型,是由一个原肾细胞特化形成的“H”型管(图7-4),伸向体后的2条纵排泄管,位于侧线内(图7-11)。
4.神经系统简单,咽部有一围咽神经环,由此向前向后各伸出6条神经(图7-12)。向后的神经中,以背神经和腹神经最发达,嵌在背线和腹线内(图7-11)。背侧神经和腹侧神经各一对,嵌在上皮内,各神经间有横神经连接。围咽神经环附近尚有一些神经节与之相连。各神经在尾端附近汇聚起来。蛔虫唇片上的唇乳突和雄虫泄殖孔前后的乳突都有感觉功能(图7-13)。
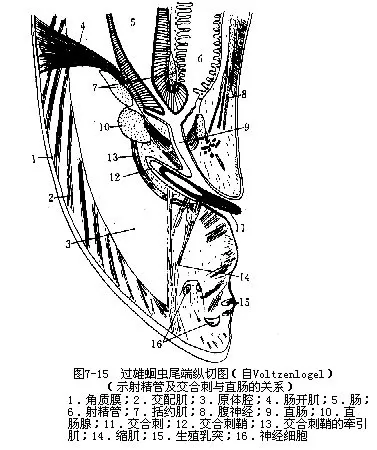
5.生殖与发育蛔虫的生殖系统发达,生殖力强。雌虫有一对细管状的卵巢、输卵管和子宫。卵巢和输卵管细,极长,前后盘曲于原体腔内,子宫较粗大,二子宫汇合成一短的阴道,以雌性生殖孔开口于体表(图7-14A)。卵巢中央有一合胞体的中轴,卵原细胞呈辐射状排列(图7-11A)。雄性为单个,也为细管状,由盘曲的精巢和输精管及较粗大的储精囊(图7-11B)和射精管组成,射精管入直肠,以泄殖孔开口于体表(图7-14B)。在泄殖腔背侧,形成一对交合刺囊,囊内各有一条交合刺(图7-15)。交配时,二交合刺伸出,可撑开雌性生殖孔,将精子经阴道排入子宫中,精子与卵在子宫远端部受精。受精卵充满子宫,有人估计约有2000万粒。一条雌蛔虫每日产卵约20万粒,生殖力惊人。受精卵呈椭圆形,大小为45μm~75μm×35μm~50μm,外被一较厚的卵壳,壳面有一层凹凸不平的蛋白质膜,可保持水分,防止卵干燥。未受精卵为长椭圆形,大小为89μm~93μm×38μm~45μm,卵壳较薄,蛋白质膜的凹凸较浅。



蛔虫为直接发育。受精卵产出后,在潮湿环境和适宜温度(20℃~24℃)下开始发育,卵裂属不典型的螺旋式,约经2周,卵内即发育成幼虫,再过1周,幼虫脱皮1次,才成为感染性虫卵,此种卵对温度及化学药物等抵抗力很强,在土壤中可生活4~5年之久。感染性虫卵被人误食,在十二指肠内孵化,数小时后幼虫即破壳外出,长仅200μm~300μm,直径10μm~15μm。幼虫穿肠壁进入血液或淋巴中,经门静脉或胸管入心脏,再到肺中。在肺泡内生长发育,脱皮2次,此时长可达1mm~2mm,后沿气管至咽,再经食道、胃到达小肠,再脱皮一次,逐渐发育为成虫。人自吞入虫卵至成虫再产卵止,大约需60~75天。蛔虫的寿命约为一年。
蛔虫生活在小肠内,其分泌物中含有消化酶抑制剂,可抑制肠内消化酶而不受侵蚀,这是寄生虫的一种适应性。人肠内有蛔虫寄生,除吸取养分外,危害并不严重。数量多时(Pyrie报道,一人肠内有蛔虫1448条),可造成肠道阻塞。成虫有迁移习性,可侵入胆管、胆囊、肝、胃等,引起不同症状,造成危害。国内有人发现,胆结石核心部分51.8%有蛔虫皮或虫卵(陈淑珍,1984)。幼虫可损伤肺、气管等,并可在脑、脊髓、眼球、肾等器官中停留,造成严重病状。
三、线虫动物门的分类
线虫有15000多种,一般分为2纲。
无尾感器纲(Aphasmida):无尾感器;排泄器官退化或无;雄虫只有一交合刺,多数营自由生活。
嘴刺目(Enoplida):咽分前后二部分,前部狭、肌肉性;后部宽,为腺体。如旋毛虫、人鞭虫(Trichuris trichiura)等。
色矛目(Chromadorida):咽分3部分,前部呈长纺锤状,中部狭,后部膨大呈球形。如扭曲线虫(Plectus parietinus)(图7-16)。
尾感器纲(Phasmida):有尾感器;排泄器官为成对的纵管;雄虫具一对交合刺;大多数营寄生生活。
小杆目(Rhabditida):咽分三部;多营自由生活,如小杆线虫。
蛔虫目(Ascaridida):咽呈长筒状,口周围有乳突。如人蛲虫、人蛔虫等。蛔虫是世界性肠道寄生线虫,种类很多。寄生在人和猪小肠中的蛔虫(Ascaris);寄生在猫、狗、狐、狼等食肉类动物的弓蛔虫(Toxocara);寄生在鸡等家禽的禽蛔虫(Ascaridia);寄生在驴马的副蛔虫(Parascar-is);寄生在蛙、蛇等两栖爬行动物的蛇蛔虫(Ophidascaris);多宫蛔虫(Polydelphis)寄生在爬行动物体内。
圆线虫目(Strongylida):咽球形或筒状;口周围无乳突。如十二指肠钩虫(Ancylostoma duo-denale)、美洲板口钩虫(Necator americanus)、粪类圆线虫(Strongyloides stercoralis)。
旋尾目(Spirurida):咽分二部分;前部肌肉性,后部为腺体。如斑氏丝虫(Wuchereria ban-crofti)、马来丝虫(Brugia malayi)。
垫刃目(Tylenchida):有口针;咽分三部分;体小。如小麦线虫(Anguina tritici)。
四、几种重要的习见线虫
人鞭虫Trichuris trichiura(Linne)(图7-17)虫体后部粗,前部3/5细长如鞭,以此钻入肠壁内。雌虫长35mm~50mm,雄虫30mm~45mm。寄生在人的盲肠和阑尾。卵在外界发育成感染性卵,在人小肠内孵化出幼虫,最后到盲肠中,经3个月才成熟。

旋毛虫Trichinella spiralis(Owen)(图7-18)成虫体小,向前端渐细。雌虫长3mm~4mm,雄虫不及2mm。人、猪、鼠为其宿主,寄生在十二指肠及空肠前部,成虫附着肠壁。雌雄交配后胎生幼虫,长仅100μm,经血液、淋巴,分布到身体各处,只有在横纹肌中才可继续发育生长。虫体卷曲,迅速增长,一般长1mm,可形成一囊胞,直径为250μm~500μm左右,内含1~2条幼虫。经6~7个月后,囊胞开始钙化(图7-18C),幼虫在内生活可达数十年。成熟囊包被宿主吞食而被感染。幼虫脱皮4次,发育为成虫。
小杆线虫Rhabditis maupasi(图7-19),本属所有种类体都极小,生活在土壤中,以腐败有机质为食,组成土壤和腐生性线虫种群。某些种类可发育成雄性先成熟的雌雄同体(protandrous hermaphrodite),行自体受精。但一般的种类为雌雄异体,如R.maupasi。虫体前端为口,具3片唇,背唇1片,侧腹唇2片,肛门在体后端腹侧。雌虫后端长且尖,雄虫后端具盘形交合伞,雌性生殖孔在体中部腹侧中线上。
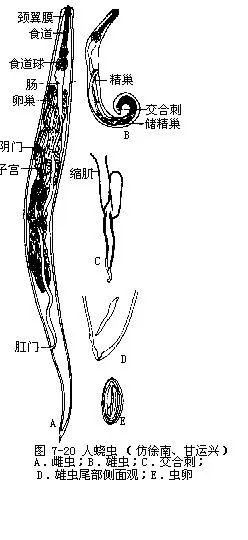
人蛲虫Enterubius vermicularis(Linne)(图7-20)成虫体细小,乳白色,似白线头状,前端具翼膜。雌虫长9mm~12mm,雄虫2mm~5mm。寄生在人的盲肠、结肠、直肠等部,虫体前端钻入肠粘膜,吸取营养。蛲虫为直接感染,儿童感染率特别高,雌虫在午夜时爬出肛门产卵,致使肛门奇痒,影响睡眠。雌虫在宿主体内生活期一般为2个月左右。公元前90年,我国《史记》中就有蛲虫的记载。


人蛔虫 Ascaris lumbricoides Linne(见图7-6)。
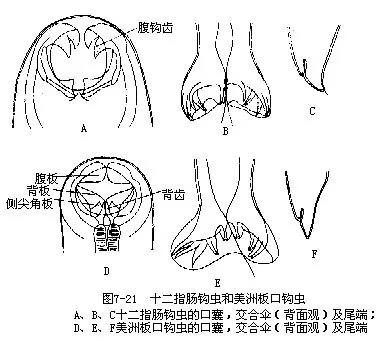
十二指肠钩虫 Ancylostoma duodenale Dubini于1843年在意大利发现,寄生在人的小肠内,大多生活1年左右,也有生活5~6年以上者。虫体小,雌虫长10mm~13mm,雄虫8mm~11mm;口囊发达,腹侧有内外2对钩齿,背侧有一对三角形齿板(图7-21A);雄虫尾端具交合伞(copulatory bursa),其背肋小枝有3个分叉(图7-21B)。钩虫以口囊吸附肠壁,摄取肠粘膜及血液为食,可使人便血、贫血、肠溃疡等,危害严重。雌虫每日排卵在2万个以上,卵在潮湿土壤中发育,经杆状蚴(rhabditiform larva)及丝状蚴(filariform larva)两期幼虫,脱2次皮。丝状蚴直接钻入人体,经血液或淋巴,过心、肺,再由气管到咽,后入胃抵肠,脱皮,吸附于肠壁,经3~4周,再脱皮,发育为成虫。钩虫分布广,属温带型,华中、华南、四川等省较为严重。
美洲板口钩虫 Necator americanus(Stile)此虫与十二指肠钩虫在形态上相似,但体较小,口囊内无钩齿,在腹侧有一对半月形齿板(图7-21D),雄虫交合伞的背肋小枝有2个分叉(图7-21E)。美洲板口钩虫属热带型,主要分布于热带、亚热带地区,我国南方多于北方。
粪类圆线虫 Strongyloides stercoralis(图7-22)成虫长2mm,生活在土壤中,环境条件恶化时,则产生感染性幼虫,幼虫穿过人的皮肤入血液中,经心、肺入肠,发育为成虫。雄虫随粪便排出,雌虫寄生在肠壁内,行孤雌生殖。产卵后,在肠内孵出幼虫,部分随粪便排出,发育成自由生活的成虫。

斑氏丝虫 Wuchereria bancrofti(Cobbold)(图7-23)寄生在人的淋巴系统,雌虫长约75mm,雄虫40mm许,可引起组织增生,使下肢、阴囊等处畸形发展,形成“象皮病”。雌雄虫交配,胎生幼虫称微丝蚴(microfilaria)。体弯曲,长200μm~300μm,体外有一鞘膜,内充满细胞核(图7-23A)。微丝蚴在人体内可生活2周以上,白天在内脏血液中,夜间则移至体表血液内。按蚊(Anopheles)及库蚊(Culex)等为其中间宿主,在蚊体内约经10~17天即可发育成感染期微丝蚴,再传给健康人。尚有马来丝虫 Brugia malayi(Brug),体较上种小,分布也不如上种广。我国山东、河南、东南沿海各省、西南各省、湖南等省都有丝虫病流行。

小麦线虫Anguina tritici(Steinbuch)(图7-24)寄生在小麦上的一种植物线虫,成虫体小,长仅3mm~4mm,雌虫向腹侧弯曲盘绕,体较雄虫粗大。寄生在小麦麦穗上,使麦粒形成虫瘿,一个虫瘿内有数千条小麦线虫的幼虫。虫瘿混在麦粒中播入土内,幼虫外出,后侵入麦苗,先在叶腋间聚集为害,使小麦发育不良,严重时不能抽穗或死亡。当小麦抽穗时,即侵入子房,迅速发育长大为成虫,子房即变成虫瘿,雌雄虫在内交配产卵,每一雌虫可产卵2000个左右。卵在虫瘿内孵出幼虫,脱皮2次,即进入休眠期。在干燥条件下,幼虫在虫瘿内可生活10年以上。次年虫瘿随小麦播入土壤中,再侵害小麦植株,有小麦线虫寄生的小麦,会严重减产。

第二节 轮虫动物门(Rotifera)
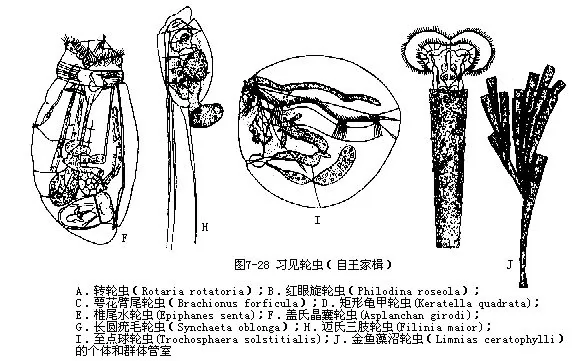
轮虫体微小,与原生动物大小相似,一般种类为100μm~500μm,最小的只有40μm左右,最大的可达4mm。轮虫为淡水浮游动物的主要类群之一,已发现2000多种,大部生活在淡水中,在湖沼、河流、水库,池塘等均有分布,咸水及海洋中种类较少。淡水轮虫多生活在浅水水域,以底栖为主,也有浮游种类。我国已报道252种。大多数轮虫分布广泛,为世界性种类,从浅水到深水都能生存。绝大多数轮虫为单体,少数为群体(簇轮虫Floscularia、巨冠轮虫Sinantherina、细簇轮虫Ptygura、沼轮虫 Limnias等,都属于簇轮虫科Flosculariidae)。轮虫以细菌、原生动物、藻类、轮虫、有机质碎屑等为食,在微酸性和微碱性水体中生活,大多数轮虫具有高度的生态耐性,是广酸碱性种类。
轮虫的身体多为纵长形,一般分为头、躯干和尾三部分。头部较宽,具有由1~2圈纤毛组成的头冠(corona),这是轮虫的形态特征之一。头冠上的纤毛不停地摆动,有游泳和摄食功能。有些种类头冠上的纤毛特化成粗壮的刚毛,有感觉作用。头冠的形式不一,有些种类的头冠上半部完全裂开,形成2个纤毛轮器(trochus),纤毛摆动,形似车轮,故称轮虫(图7-25)。
轮虫体被角质膜,常在躯干部增厚,称为兜甲(lorica),其上往往形成刺或棘。一些部位的角质膜因硬化程度不同而形成环形的折痕,形似体节。当身体收缩时,前后端的节可向中部缩入,如套筒状。角质膜下为合胞体的上皮,环肌及纵肌成束,发达,因此轮虫伸缩自如。轮虫为原体腔,腔内充满体腔液。
尾部又称足,为躯干部向后逐渐变细形成,长筒状,少数浮游种类无足。足内具足腺,借其分泌物可粘附于其他物体上。足末端一般有一对趾(toe),有的种类有3或4趾,趾在爬行时有固着于底层的作用。
轮虫的消化管分为口、咽、胃、肠、肛门等。口位头部腹面,咽部特别膨大,肌肉发达,又称咀嚼囊(mastax)。咽内具有咀嚼器(tro-phi)(图7-25),这也是轮虫的特征之一。咀嚼器形式多样,为分类的重要依据。它是由角质膜硬化形成的多块坚硬的咀嚼板构成,主要有7块:单块的基骨(fulcrum)及成对的枝骨(ramus)、爪骨(un-chus)及槌骨(manubrium)(图7-26)。咀嚼器不停地运动,可磨碎食物。咽侧有一对或数个唾液腺。咽由管状食道通入膨大的胃,胃囊状,内壁具纤毛。一般胃前有一对胃腺,可分泌酶,有开口通入胃。胃是消化和吸收的主要部位。肠管状,较短,连于泄殖腔,以泄殖孔开口于躯干与愿交界处的背侧(图7-25)。

排泄器官为一对由排泄管和焰球(flame bulb)组成的原肾管,位体的两侧(图7-25),为合胞体细胞衍生而成,细胞核位排泄管的管壁中,这点与涡虫显然不同。2排泄管通入膀胱,与肠汇合入泄殖腔,由泄殖孔开口于体外。

轮虫为雌雄异体,但雄性个体不常见,且体小,仅有雌的1/8~1/3,寿命短,体内只有一精巢一输精管及阴茎,其他器官均退化。有些种类(蛭态目)从未发现过雄性个体。雌轮虫的卵巢、输卵管、卵黄腺等一般都是单个的,少数种类左右成对(双巢目)。
轮虫在环境条件良好时营孤雌生殖,雌轮虫产的卵不需受精,称非需精卵(amictic egg),其染色体为双倍体(diploid),即卵成熟时不经减数分裂,此卵可直接发育成雌性个体,称为非混交雌体(amic-tic female)。经多代孤雌生殖,当环境条件恶化时,孤雌生殖产生混交雌体(mictic female)。混交雌体产生的卵成熟时经减数分裂,卵的染色体为单倍体(haploid),只有这种卵才能受精,故称为需精卵(mictic egg)(图7-27)。轮虫受精作用在体内进行,一般是雄轮虫以阴茎刺破雌轮虫的体壁,将精子输入原体腔内。卵受精后,分泌一层较厚的卵壳,可以抵御不良环境,称休眠卵(resting egg)。当外界条件好转时,发育成非混交雌体,继续进行孤雌生殖。如需精卵未能受精,则发育成雄性个体。轮虫的非混交体每年出现数10代,而混交雌体一年只出现1或2代。食物和种群密度都影响产生混交雌体的比例,轮虫摄入维生素E及种群密集情况下,可产生混交雌体。轮虫这种周期性孤雌生殖与环境有着密切的关系,是对环境周期性变化的一种适应性。
当轮虫生活的水体干枯时,有些种类仍能生存。轮虫的身体失去大部分水分,高度卷缩,进入假死状态,耐干燥能力极强,抵抗干燥的环境几个月到几年。再入水后,即能复活。这种状态维持生存,称为隐生(cryptobiosis)。
轮虫的神经系统主要由位于咽背侧的双叶状脑神经节及伸向体后的2条腹神经组成,脑神经节向体前端和背侧发出许多神经。轮虫的神经系统与涡虫的极为相似。感觉器官位头部,有头冠上的感觉毛,眼点1~2个(一般为一个),一条背触手及2条侧触手,触手为短棒状突起,其末端具感觉毛。
轮虫的各器官组织的结构均为合胞体,且各部分含有的细胞核数目是恒定的。如水轮虫(Epiphanes senta)的上皮层含有280个细胞核,食道含15个,胃含39个,肠含14个,原肾管(一条)含14个,脑含183个,周围神经含63个,细胞核总数为959个。这表明轮虫的发育有一个显著的固定形式。轮虫自卵孵出后,细胞核即不再分裂。身体部分受损,也不能再生。

第三节 腹毛动物门(Gastrotricha)
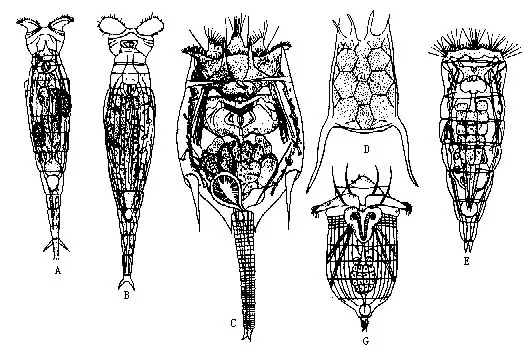
腹毛动物是一类身体微小的水生动物,多数生活在海洋中,少数在淡水,已知200多种。腹毛动物体呈圆筒状,长0.1mm~1.5mm,但一般都小于0.6mm。体被角质膜,背面略隆,其上常见有刚毛、鳞片、棘等(图7-29A、B),有感觉作用。腹面平,具有若干纵行或横排纤毛,故称腹毛动物。借纤毛摆动可游泳或爬行,有些种类纤毛不发达(如 Proichthydium、Setopus、Lepido-dasys等)。

腹毛动物的上皮为合胞体,其下纵肌成束;原体腔。消化管完整,口位体前端,周围有长纤毛束或棘毛(bristle)丛,司感觉。肛门在体后端腹面(图7-29C)。体末分2叉,每叉的端部有粘腺开口,粘腺的分泌物有粘附作用。排泄器官为一对具焰球的原肾管,位消化管中部两侧,排泄孔开口于腹面中央(图7-29C)。有些种类无原肾管,具腹腺。一对脑神经节位咽的前端,一对侧神经与之相连。绝大多数(海产种类)为雌雄同体,有些种类的精巢退化(如 Chaetonotoida),只有雌性个体出现,故营孤雌生殖。卵巢一个或一对,有子宫、纳精囊(seminal receptacle)、交配囊和雌性生殖孔。精巢一个或一对,有输精管、雄性生殖孔,有的种类输精管通入交配囊。卵具厚壳,直接发育。
淡水中习见种类如鼬虫(Chaetonotus),海产的如头趾虫(Cephalodasys)、大趾虫(Macro-desys)及尾趾虫(Urodasys)等(图7-29)。
第四节 原腔动物的系统发生
线虫动物具有特殊的排泄管,无纤毛,特殊的纵肌层,线形生殖系统。这些结构特点与原腔动物中其他类群显然不同,它们是动物演化上的一个杜塞分枝。腹毛动物体表具角质膜,原体腔,尾具粘腺。这些与自由生活的线虫相似。另一方面,体表有纤毛,具焰球的原肾管,双腹式神经,大多数种类为雌雄同体。这些特点似涡虫纲。因此通过腹毛动物说明线虫动物和涡虫纲在演化上有着一定的类缘关系。
轮虫的构造和胚胎发育与涡虫纲相似。许多轮虫体形较扁,具纤毛的头冠显著偏向腹面。具焰球的原肾管与涡虫纲单肠目动物相同。雌雄异体,具卵黄腺,胚胎发育中早期卵裂属螺旋形,双腹式神经。这些特点说明轮虫可能由涡虫纲演化而来。但轮虫为发育完善的消化管,具特殊的咀嚼器,各组织器官为合胞体,且细胞核的数目恒定。这些显然又不同于涡虫纲。总之,轮虫动物与涡虫纲在演化上有着较为接近的类缘关系。轮虫具足腺,有纤毛,具焰球的原肾管,与腹毛动物接近。因此有的分类系统将这二类动物列为担轮动物(Trochelminthes)。
附一:棘头动物门(Acanthocephala)
棘头动物全部为寄生种类,寄生在脊椎动物的肠管内,有500多种。体呈圆筒形或稍扁平,大小差异很大,长1cm~65cm不等,一般长25cm以下。体前端有一能伸缩的吻(proboscis),可缩入吻鞘内,吻上具有许多倒钩(图7-30),为附着器官,钻入宿主肠内后可钩挂在肠壁上,故称棘头虫。体表具角质膜,上皮为合胞体,其内贯穿着复杂的腔隙系统(lacunar system),是储存营养之处。上皮内为环肌和纵肌组成的肌层,这点与线虫动物等显然不同。原体腔,无消化管,以体表吸收宿主肠内的营养物质。排泄器官若有时为一对具焰球的纵行原肾管,位体的两侧,汇合后与输精管或子宫相通,由生殖孔通体外。位吻鞘处有一神经节,由此伸出神经至身体各部。棘头虫雌雄异体,其生殖器官结构特异,雄虫有精巢一对及输精管、阴茎、雄性生殖孔;雌虫有卵巢一个或一对,体后有一特殊的子宫钟(uterine bell),为一肌肉性漏斗形管,上有2对孔,前一对通原体腔,后一对孔扁,通阴道。未成熟卵不能通过后一对孔,而由前一对孔重新回到原体腔内。成熟的卵才可通过后一对孔,经阴道由雌性生殖孔排出体外。卵被中间宿主昆虫、甲壳类等吞食,在其体内发育,当终末宿主吞食中间宿主时,即被感染,在肠管内发育为成虫。
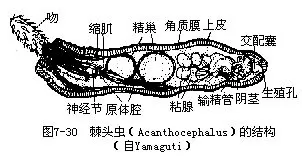
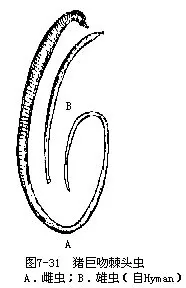
常见的如寄生在猪小肠内的猪巨吻棘头虫(Macracanthorhychus hirudinaceus)(图7-31),为最大的一种棘头虫,雌虫长65cm,雄虫15cm,幼虫寄生在金龟子的幼虫蛴螬体内,猪吞食蛴螬而被感染,以吻固着于肠壁上,发育为成虫。棘头虫寄生,影响猪的生长发育,严重时可使猪死亡;鱼棘头虫(Echinorhychus proteus)寄生在淡水鱼体内。世界上已记录棘头虫有 1000多种,我国已知寄生在鱼类、鸟类及哺乳类体内的棘头虫有135种,隶属于18科44属,分布在福建、台湾、黑龙江、湖北、四川等14省区。
棘头动物具原肾管,肌层由环肌与纵肌构成,这与涡虫纲近似,但棘头动物具吻,有复杂的腔隙系统,无消化管,特异的生殖器官等特点,与原腔动物中其他类群显然不同,其演化地位难以确定。
附二:线形动物门(Nematomorpha)
线形动物体呈线形,粗细一致,细长,自0.5m~1m或更长,直径只有1mm~3mm,已知100多种。体被较硬的角质膜,其下为上皮,细胞界限清楚;上皮内为纵肌;消化管在成体及幼体均退化,常常无口,不能摄食,以体壁吸收宿主的营养物质。肠壁为一单层细胞的上皮,在组织学上与昆虫的马氏管相似,可能有排泄功能;消化管后端与生殖导管相连,形成泄殖腔,具有角质膜衬里;排泄孔开口于体后端腹面,无排泄器官;原体腔内充满间质(mesenchyme)(图7-33);神经与上皮相连,体前端有一神经环,向后伸出一腹神经;雌雄异体,雄体较小,向腹侧卷曲。

线形动物的成虫生活在河流、池塘等淡水中,多在春季雌雄交配产卵,卵粘成索状。孵出的幼虫具有能伸缩的有刺的吻,借以运动,在水底生活。幼虫钻入宿主体内或被宿主吞食,即营寄生生活。寄生在昆虫类的螳螂、蝗虫、龙虱等体内,逐渐发育为成虫,离开宿主,在水中营自由生活。如宿主身体过小,幼虫即停止发育,当这个宿主被更大的宿主吞食,再在新宿主体内继续发育。如此出现了更换宿主的现象。
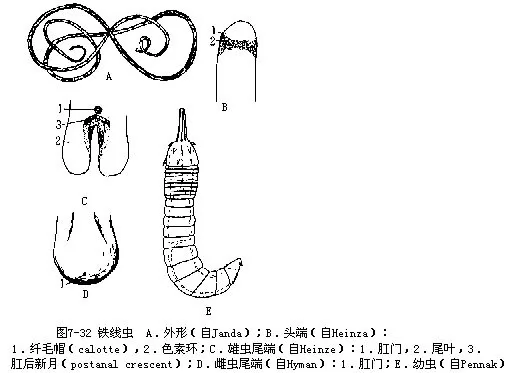
习见种类如铁线虫(Gordius aquaticus)(图7-32),成虫长10cm~30cm,直径0.3mm~2.5mm,自由生活,像一团生锈的铁丝状,雄虫体末端分二叉。离开水时,耐干旱,再遇到水则恢复运动。寄生在蝗虫,螳螂体内,当这些昆虫落入水中,成虫即离开宿主,营自由生活。
线形动物的体形和某些结构似线虫动物,但体无侧线,原体腔中充满间质,消化管退化,无排泄器官,神经结构特殊,故其演化关系尚不明确。
复习题
1.原腔动物的主要特征是什么?哪些门动物属于原腔动物?
2.比较原腔动物中各类群动物间的异同。
3.试述人蛔虫形态结构及生活史特点,并说明它的哪些特点代表了线虫动物门的特点。
4.分析人蛔虫的生活史,说明其感染率高的主要原因。
5.比较蛲虫、钩虫、丝虫及旋毛虫的结构及生活史的异同。
6.试述寄生线虫对寄生生活方式的适应性。
7.从腹毛类的特点,说明原腔动物在系统演化中的位置。
8.试述轮虫动物的主要特征、生殖发育的特殊性及其经济意义。
9.线形动物适应生存的特点为何?
10.棘头虫类表现出适应寄生生活的特征是什么?
11.试述原腔动物与人类的关系。
本文标题:第七章 原腔动物(Protocoelomata) 附:棘头动物门、线形动物门
手机页面:http://m.dljs.net/dlsk/dongwuxue/59426.html
本文地址:http://www.dljs.net/dlsk/dongwuxue/59426.html
