第一节 棘皮动物门的特征
棘皮动物全部生活在海洋中,身体为辐射对称,且大多数为五辐射对称,但这是次生形成的,是由两侧对称体形的幼体发展而来。
棘皮动物的次生体腔发达,是由体腔囊又称肠腔囊(enterocoel)发育形成,即在原肠胚期,于原肠的背侧凸出成对的囊,以后囊脱落,形成中胚层,发育成次生体腔。
体壁由上皮和真皮组成,上皮为单层细胞,真皮包括结缔组织、肌肉层及中胚层形成的内骨骼,真皮内面为体腔上皮。内骨骼有的极微小(海参类),有的形成骨片,呈一定形式排列(海星类、蛇尾类及海百合类),也有的骨骼完全愈合成一完整的壳(海胆类)。内骨骼常突出于体表,形成棘或刺,显得皮肤很粗糙,故称棘皮动物。
棘皮动物特有的结构是水管系(water vascular system)和管足(tube foot)。这是次生体腔的一部分特化形成的一系列管道组成,有开口与外界相通,海水可进入循环。水管系包括:环管(ring canal)、辐管(radial canal)和侧管(lateral canal)(见图12-7)。侧管连于伸出体表的管足,管足有运动、呼吸及摄食功能。依管足的分布,棘皮动物的身体可以区分为10带区,有管足的带区称步带(ambulacrum),无管足的带区称间步带(interambulacrum),二者相间排列(图12-15)。
棘皮动物一般运动迟缓,故神经系统和感官不发达。雌雄异体,个体发生中有各型的幼虫(羽腕幼虫、短腕幼虫、海胆幼虫、蛇尾幼虫、樽形幼虫、耳状幼虫、五触手幼虫等)。
第二节 代表动物——海盘车
海盘车(Asterias)一般称海星,分布在世界各海区,以太平洋海域种类最多。海盘车生活在潮间带的礁岩间或海底。运动缓慢,肉食性。海盘车主要以瓣鳃类为食,对人工养殖的双壳类有一定的危害。海盘车再生力强。我国黄渤海的习见种类为罗氏海盘车(Asterias rollestoni Bell)。
(一)外部形态 海盘车体呈五角星状,由体盘及腕组成,二者界限不明显。腕一般5条,故体呈典型的五辐射对称(图12-1Aa)。
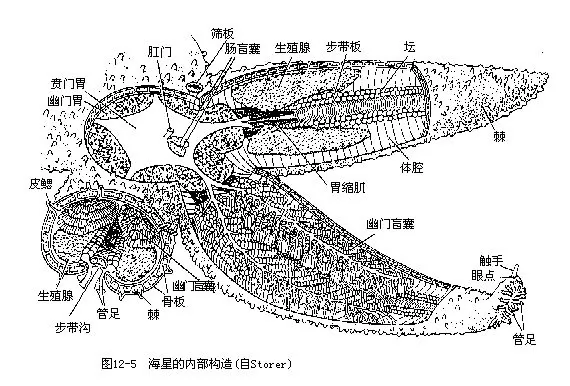
自体盘中心至腕末端的距离(以 R表示)可达100mm以上。生活时口面向下,反口面向上。口面平坦,淡黄色,体盘中央为口,周围有围口膜(peristomial membrane)。各腕腹侧中央有一条自口伸向腕端部的步带沟(ambulacral groove)(图12-1A之b),其内伸出管足2排(外观上似为4排),管足的末端有吸盘。步带沟侧缘具二、三列可动的棘。反口面略隆起,有紫红色花纹,颜色鲜艳。近体盘中央处有肛门,极小。两腕之间为间步带区,在一间步带区有一块圆形多孔的小板称筛板(madreporite),为海水出入之口。各腕的基部两侧各具一对生殖孔。腕顶端有1条触手,其下有红色眼点(图12-5)。海盘车的管足为运动器官,由于管足伸出向后撑,推身体前进。
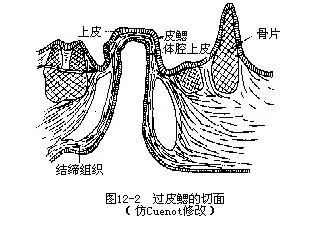
海盘车体表粗糙,许多内骨骼向外突出成棘(papilla)或刺(spine),棘间分布有叉棘(pedicel-laria)和皮鳃(papula)(图12-1B)。叉棘很小,由3个小骨片构成,一个基片(basal ossicle)和2个颚片(blade)(图12-1C)呈钳状,能活动,有清除体表污垢之功。皮鳃呈泡状,为体壁自骨片间隙外凸形成。外层为上皮,内为体腔膜(图12-2),体腔液在其内循环,有呼吸及排泄的功能。
(二)内部构造
1.体壁体表为一层柱状上皮,其外有一层薄的角质膜。上皮细胞间散布有腺细胞和神经感觉细胞,基部为一薄的基膜与真皮分开。基膜下为一神经层。真皮较厚,包括结缔组织和肌肉层。结缔组织分泌小骨片,并将它们联系在一起,成网状骨骼。肌层外为环肌纤维,内为纵肌纤维,均属平滑肌。反口面沿各腕背中线辐射伸出的纵肌束较发达。体壁最内层为体腔上皮,具纤毛(图12-3)。在肌层外侧真皮内,充满一系列空隙,有的空隙形成一环,围绕于皮鳃的基部(图12-2)。内骨骼常常突出于体表形成棘,外覆有上皮。

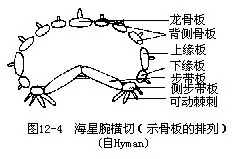
位体壁中的内骨骼,排列成窗格状,其间充满结缔组织。口面腕中央为两行不带棘的步带板(ambulacral plate),构成步带的底壁,前后步带板间有排列整齐的小孔,管足即由此孔伸出体外。步带板两侧各为一行侧步带板(adambulacral plate),其上有一细长的可活动的侧步带棘(adam-bulacral spine),有保护管足的功能。侧步带板上面为下缘板(supramarginal plate)和上缘板(in-framarginal plate)。反口面腕中央为一块龙骨板(carinal ossicle),具棘。龙骨板与缘板之间为一系列背侧板(dorsolateral plate)(图 12-4)。这些骨片每一组彼此相连,各组间有活动关节,故腕可上下灵活运动。
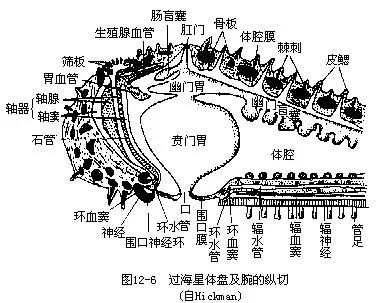
2.体腔 海盘车次生体腔发达,围绕消化管和生殖腺,直伸至腕的顶端(图12-6)。体腔的一部分形成了水管系和围血系统(perihaemal system)。体腔内充满体腔液,由于体腔上皮细胞的纤毛摆动,使体腔液流动,起到运输的作用。体腔液内有两种变形细胞,由体腔上皮产生,有吞噬作用,吞食的颗粒自皮鳃排出体外。此外尚有很少的无色小球形的体腔细胞存在。
3.消化系统 消化管短而直,自口面伸向反口面。口位于体盘正中,围口膜中央处,口周围有括约肌和辐射肌纤维。经短的食道进入宽大的充满体盘的胃。胃分为近于口面的贲门胃和近于反口面的幽门胃两部分,二者之间有一缢缩。贲门胃大,多绉褶;幽门胃小,扁平,向各腕内伸出 2支盲囊,称幽门盲囊(pyloric caeca)。能泌酶。胃后为很短的肠,末端开口为肛门(已无肛门作用)。肠尚有一分2或3支的肠盲囊(intestinal caeca)(图12-5,图12-6)。
消化管的组织似体壁,只是无骨骼。幽门盲囊的组织结构同消化道,只是上皮更厚。具纤毛的上皮细胞可分为腺细胞、贮存细胞和粘液细胞。腺细胞可泌酶,贮存细胞内充满类脂小滴、少量糖原和某些多糖——蛋白质复合物。当动物饥饿时,贮存的食物即消失。肠盲囊的上皮多皱,含有粘液细胞和腺细胞。
海盘车肉食性,以软体动物、棘皮动物、蠕虫等为食,口能扩张,能吞入较大的动物。捕食瓣鳃类时,身体作弓形隆起,以腕包住动物,管足吸在贝壳上,向左右拉开二壳,将贲门胃翻出,包住动物的肉体,先行局部消化,后将动物吞入胃内,胃缩回体内。消化主要在幽门胃中进行,已消化的营养物质为幽门盲囊吸收贮存,养分可透过盲囊入体腔液内,运送至身体各部分。
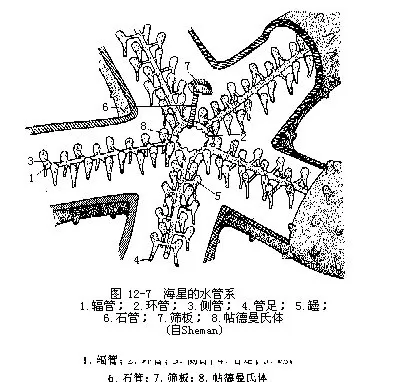
4.水管系统 为棘皮动物的特有器官,从发生上看,是次生体腔的一部分特化形成。环管位口面口的周围,自此向每个腕辐射出一条辐管,辐管两侧各伸出侧管,其端部连于管足(图12-7)。管足上部为一囊状的罎(ampulla),下有一管,末端具吸盘。管足为内外交错排列,外观上每辐管一侧尤如两排。借罎的伸缩,使管足吸附外物,可捕食,并利用腕的弯曲以管足支撑外物而推身体向前,完成运动。
环管于一间步带处向反口面伸出一石管(stone canal)(位于轴器内),管壁内有石灰质环,故较硬,管末端连于筛板,与外界相通。筛板为一圆形小骨板,其上有许多辐射排列的小沟纹,沟底有许多小孔(约200个)。环管上间步带处各具一对帖德曼氏体(Tiedmann’ s body)(图12-7),在石管连接环管处只有一个,故总共有9个。帖德曼氏体是一种特殊的小型不规则的腺体组织,内腔含有体腔细胞,可能产生变形细胞。有些种类于此位置上尚有波里氏囊(Polian vesicle)(1~5个)有调节水管系内的水压作用。
5.围血系统包括生殖窦(genital sinus)和环窦(ringsinus)两部分(图 12-8),为次生体腔的一部分(左后体腔囊的一部分分离形成)特化形成。生殖窦位反口面体盘的体壁下方,为一五边形管,向每一生殖腺伸出一分支,后膨大形成包围生殖腺的囊。环窦位口面,口的周围,环管之下。为一圆形管。环窦内有一斜行隔膜,将其分为内环窦与外环窦两部,前者小于后者。环窦向各腕伸出一条辐窦(radial sinus),其内有一垂直隔膜。
轴窦(axial sinus)为一薄壁管状的囊,位体盘有筛板的间步带,包括石管和轴腺(axial gland)两部分,二者的壁由系膜紧紧结在一起。轴窦和轴腺合成轴器(axial organ)。轴腺是由许多小血窦组成的海绵状结缔组织,外被体腔上皮。轴窦在反口面与生殖窦相通,此处还伸出一能收缩的背囊(dorsal sac),内腔有轴腺突入。轴窦在口面连于环窦。
各血管均位于各血窦内,因此称为围血系统(perihaemalsystem)。

6.血系统 此系统很退化,只有在切片上方可看清。环血管(oral haemal ring)位环窦的隔膜内,由此向各腕伸出一辐血管(radial haemal canal)(图12-8),在辐窦的隔膜中穿过,有分支至管足。自环血管向上伸出一血管丛,入轴腺内,连于位生殖窦内的反口环血管(aboral haemal ring),此处有分支到生殖腺,称生殖血管,位生殖窦的分支内。
海胆类和海参类的血系统(haemal system)较明显。
7.呼吸与排泄 气体交换主要通过皮鳃进行,管足也起着一定作用。代谢产物由体腔液中的变形细胞吞食,经皮鳃排出,主要是氨和尿素。
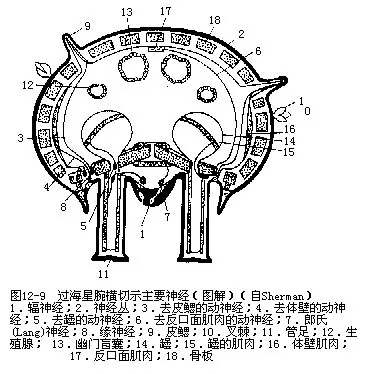
8.神经系统和感觉器官 海盘车的神经系统包括3个有联系的系统。口或外神经系(oral or ectoneural system),为神经环、辐神经及神经丛组成(图12-9),正位于上皮之下。神经环呈五角形,位围口膜周围,环窦下面,有神经通至围口膜及食道。神经环向各腕伸出一条辐神经,位步带沟底,各辐窦之下,终止于端触手的反口面一侧,横切面呈“V”形,其外侧与上皮相连(图12-9)。神经丛位体壁上皮下,辐神经与之相连续。外神经系司感觉。下神经系(hyponeural system)位环窦壁的侧面部分,为一神经环及5条辐神经组成,位体腔上皮下,与之相连,由真皮的一薄层结缔组织与外神经系隔开。下神经系正位于外神经系之上,有分支至腕、管足、叉棘等器官的肌肉,司运动。反口神经系(aboral system)位反口面,由体腔上皮产生,这点很特殊。此神经系无神经环,只有5条辐神经,在海星类不显著,海百合类明显,司运动。
海盘车的感觉器官不发达,在上皮间散布着许多呈棱形的神经感觉细胞(neurosensory cell),可能有触觉器和化学感觉器两种功能,在管足的吸盘处数目最多,在棘和叉棘基部的上皮处,每mm2可达70 000个。在各腕的顶端触手的基部口面有一眼点,由一群感光细胞和色素细胞构成,可感光。海盘车口周围的管足有嗅觉功能。
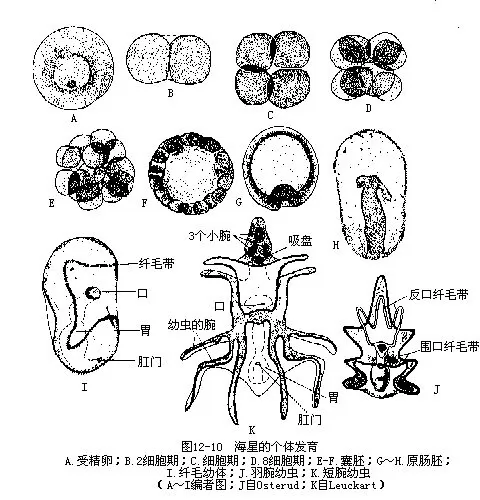
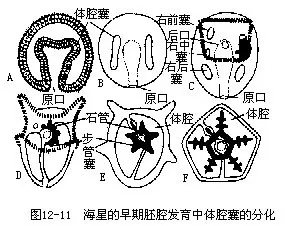
9.生殖和个体发育海盘车在各腕内基部两侧反口面间步带处,共有5对生殖腺,各附着在近腕间隔处。其附着处即为生殖孔,也为5对,极小。生殖导管很短。非生殖季节期,生殖腺不发达,当生殖季节时,可充满腕内,伸至腕端(图12-5,图12-9)。海盘车雌雄异体,精卵在海水中受精,经完全均等卵裂,有腔囊胚,以内陷法形成原肠胚(图 12-10 A~H)。以体腔囊法形成中胚层和体腔,这与其他的无脊椎动物完全不同。胚胎继续发育,逐渐延长,左右体腔囊也随之延长,且分裂为前、中、后三部分。左侧各部发育良好,右侧部分则多退化消失。前中部分相连,有一共同通道,开口于背侧。一对后体腔囊愈合,发育成动物成体的次生体腔;左后体腔囊的一部分分离,形成围血系统;左中体腔囊演变成水管系;左前体腔囊分化成轴窦;前中体腔囊的共同开口成筛板。右前体腔囊和右中体腔囊在胚胎发育中逐渐退化消失(图12-11)。胚胎延长,原口移至腹面成为肛门,在胚的另端形成口,这是后口动物的特征。消化管已形成,呈“U”形。体表出现纤毛,可在水中游泳并摄食,称纤毛幼体(图12-10I)。此后继续发育,椭圆形的幼体产生腕,体表纤毛退化,只在腕周围形成纤毛带,此为羽腕幼虫(Bipinnaria)(图12-10J)。后在背面基部产生二突起,进入短腕幼虫(Branchialaria)期(图12-10 K),体两侧对称,前端具有3个小腕及一个吸盘。短腕幼虫在海水中游泳一时期后,沉入水底,营固着生活,进入变态期。幼虫的口、食管、肛门、肠等退化,只消化管中间一段发达。随着水管系的发育,逐渐由两侧对称变成辐射对称,原来胚的左侧成为身体的口面,右侧成为反口面,退化的器官重新形成。变态结束,最后发育成为轴射对称的小海星。
海盘车有很强的再生能力,腕、体盘受损或自切,均能再生。单独的腕不能再生出完整的身本(Linckia属除外)。
三节 棘皮动物的分类
棘皮动物为一古老的类群,始于古生代寒武纪,到志留纪、石炭纪、泥盆纪最繁盛。生存种类共约6000种,包括海星、海参、海胆、蛇尾、海百合等,我国已记录 300多种。根据动物的体形、有无柄和腕、筛板的位置以及管足的结构等,分为2亚门,5纲;尚有4纲全为化石种类。

一、有柄亚门(Pelmatozoa)
幼体终生具柄,固着生活。口面向上,反口面向下,肛门完整的壳;主要神经系统在反口面。生存种类约650种,只有1纲,另4纲均为化石种。
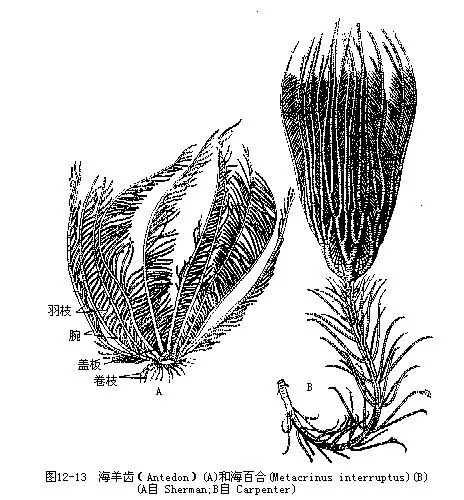
(一)海百合纲(Crinoidea)海百合类多生活在深海中,底栖,营固着生活。一类终生具柄,称海百合类(stalked crinoids),一类成体无柄,为海羊齿类(comatulids)。海羊齿类多栖息于沿岸浅海岩礁底,可附着外物或自由游泳生活。
海百合体分根、茎、冠三部(图12-12)。茎一般称柄,由许多骨板构成,其上常有分支的附支,称为根卷支(radiculus),有附着作用。冠由萼(即体盘)和腕构成,萼呈杯状或圆锥状,背侧由石灰质骨板组成,具口、肛门、步带沟。步带沟内生触手,无运动功能,可捕食。海羊齿的萼称体盘,腕原始为5个,但由于一再分支而成多个。腕由多数腕板构成,两侧具有许多羽支(pinnule)。消化管完整,主要以浮游动物为食。生殖腺位生殖羽支(genital pinnule)(羽支的一种)内,个体发育中有桶形的樽形幼虫(doliolaria)。海百合类再生能力极强,常将腕甚至萼等一起断落。后再生。
海百合(Metaerinus)具长柄(图 12-13 B),卷枝(cirrus)多,我国海南有分布,生活在水深200m处。海羊齿(Antedon)(图12-13A)无柄,营自由生活。锯羽丽海羊齿(Compsometra serrata)青岛胶州湾、台湾有分布。
二、游移亚门(Eleutherzoa)
无柄,自由生活。口面向下,口位口面或体前端,肛门位反口面或体后端。骨骼发达或不发达;主要神经系统在口面。分4纲。
(二)海星纲(Asteroidea)体扁平,多为五辐射对称,体盘和腕分界不明显。生活时口面向下,反口面向上。腕腹侧具步带沟,沟内伸出管足。内骨骼的骨板以结缔组织相连,柔韧可曲。体表具棘和叉棘,为骨骼的突起。从骨板间突出的膜质泡状突起,外覆上皮,内衬体腔上皮,其内腔连于次生体腔,称为皮鳃(papula),有呼吸和使代谢产物扩散到外界的作用。水管系发达。个体发育中经羽腕幼虫和短腕幼虫。本纲约有1600种。
显带目(Phaneroznia):腕的上、下缘板大而显著;管足无吸盘,叉棘无柄。砂海星(Luidea)口面与反口面间的上缘板和下缘板大而显著,如一镶边状(图 12-14 A)。
有棘目(Spinulosa):上下缘板不显著;管足有吸盘,很少有叉棘。海燕(Asterina pectinifera)体形近五角形,体表具颗粒状小棘。太阳海星(Solaster)腕10余条(图 12-14 B、C)。
钳棘目(Forcipulata):上下缘板不显著;管足有吸盘;具叉棘为最大特点。海盘车(Asteriasrollestoni),鸡爪海星(Henricia leviuscula)腕长而圆,末端细(图12-14 D、E),此2种均为华北沿海习见种类。
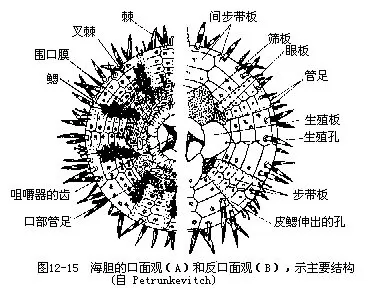
(三)海胆纲(Echinoidea) 体呈球形,盘形或心脏形,无腕。内骨骼互相愈合,形成一坚固的壳。壳板分三部:第一部最大,由20行多角形骨板,排列成10带区,5个具管足的步带区和5个无管足的间步带区,二者相间排列。各骨板上均有疣突和可动的长棘。第二部称顶系,位反口面中央,由围肛部(periproct)和5个生殖板,5个眼板(ocular plate)组成。生殖板上各有一生殖孔,有一块生殖板多孔,形状特异,兼作筛板的作用。眼板上各有一眼孔,辐水管末端自孔伸出,为感觉器。围肛部上有肛门。第三部为围口部,位口面,有5对口板,排列规则,各口板上有一管足。口周围有5对分支的鳃,为呼吸器官(图12-15)。

海胆的壳上生有疣突及可动的细长棘,有的棘很粗。多数种类口内具结构复杂的咀嚼器,称亚里斯多德提灯(Aristotle’s lantern),其上具齿,可咀嚼食物。消化管长管状,盘曲于体内,以藻类、水螅、蠕虫等为食。
海胆类多为雌雄异体,个体发生中经海胆幼虫,后变态成幼海胆,经1~2年才达性成熟。
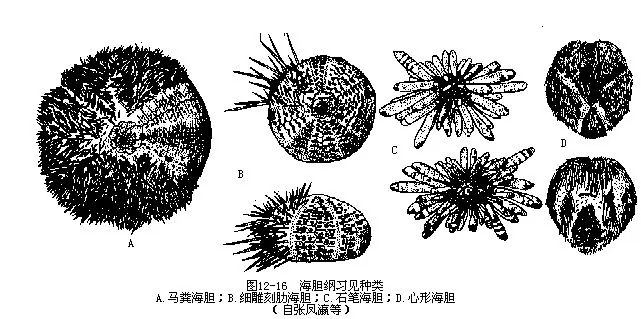
海胆有800多种,习见种类有马粪海胆(Hemicentrotus pulcherrimus)壳半球形,褐色,棘短而多,状如马粪;细雕刻肋海胆(Temnopleurus toreumaticus)壳较低平,棘大而长;石笔海胆(Heterocentrotus mammillatus)棘粗大如石笔状(图12-16A~C),我国西沙群岛有分布。这些海胆都略呈半球形,辐射对称,故属正形目(Centrechinoidea)。心形海胆(Echinocardium corda-tum)(图12-16D)壳似心脏形,薄而脆,口和肛门均位口面,属心形目(Spatangoidea)。
(四)蛇尾纲(Ophiuroidea) 体扁平,星状,体盘小,腕细长,二者分界明显。腕内中央有一系列腕椎骨(vertebra),骨间有可动关节,肌肉发达。腕只能作水平屈曲运动,很灵活。腕上常被有明显的鳞片,无步带沟。管足退化,呈触手状,无运动功能。每一腕节由4块腕板组成,上下左右各一,侧腕板上生有腕棘,侧腕板间有2列触手孔,触手自此伸出。触手孔边有触手鳞。消化管退化,无肠,无肛门。以藻类、有孔虫、有机质碎屑为食,也食多毛类、甲壳类等小动物。个体发生中经蛇尾幼体(ophiopluteus),有少数种类雌雄同体,胎生。
蛇尾类约有200种,腕分支的为蔓蛇尾目(Euryalae),如筐蛇尾(Gorgonocephalus),腕分支,筛板多个,产海南东部海区。海盘(Astrodendrum)腕分支,产黄海。腕不分支的为蛇尾目(Ophiurae),如真蛇尾(Ophiura)。滩栖阳遂足(Amphiura vadicola)腕极长,可达180mm以上,青岛有分布。刺蛇尾(Ophiothrix fragilis)(图12-17)

(五)海参纲(Holothuroidea) 体呈蠕虫状,两侧对称,背腹略扁,具管足,背侧常有疣足(pa-pillae)(一种变形的管足),无吸盘或肉刺(warts)。口位体前端,周围有触手,其形状与数目因种类不同而异,肛门位体末。内骨骼为极微小的小骨片,形状规则。消化道长管状,在体内回折,末端膨大成泄殖腔。由此分出一对分支的树状结构,称呼吸树或水肺,为海参特有的呼吸器官。受刺激时,可从肛门射出,抵抗和缠绕敌害,以后可以再生。另有许多盲管状的居维尔氏器(Cuvierian organ),有排泄功能。围绕食管有石灰环(calcareous ring)特有结构,由5个辐片和5个间辐片构成。各辐片前端有孔或凹痕,辐水管和辐神经由此通过。筛板退化,位体内。

海参在海底匍匐,食物为混在泥沙内的有机质碎片、藻类及原生动物等,摄食时,连同泥沙一同吞入。个体发育中经耳状幼体(auricularia)(图12-18)、樽形幼虫(doliolaria)和五触手幼虫(pentactula),变态成幼参。
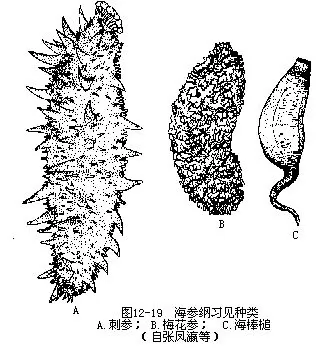
海参从沿海到万米深的海沟都有分布。共约1000多种。我国华北沿海习见种类有:刺参(Stichopus japonicus)为大型食用参,体壁厚,肉刺大,含蛋白质高,味鲜美。梅花参(Thelenota ananas)体长700mm左右,大的可达1m,是海参中最大的种类,体上肉刺基部相连,成梅花瓣状,为我国南海产的食用参中最好的一种。以上海参的触手不分支,末端有小突起,属楯手目(Aspidochirota)。海棒槌(Paracaudina chilensis var.ransonnettii)又称海老鼠,体呈纺锤形,后端延长成尾状,体表光滑。管足和肉刺均退化,属芋参目(Molpadonia)(图12-19)。
第四节 棘皮动物的经济意义
棘皮动物中有些种类对人类有益,少数有害。海参类中有40多种可供食用,它们含蛋白质高,营养丰富,是优良的滋补品。我国的刺参、梅花参等为常见的食用参。海参又可入药,有益气补阴、生肌止血之功。我国已成功地进行了人工繁殖饲养刺参,辽宁海洋水产研究所获得了幼参放流1.5周年成活率为55.3%的良好结果。海胆的卵可食用,据记载我国明朝已有了以海胆生殖腺制酱的应用。海胆卵为发育生物学的良好实验材料。海胆壳入药,可软坚散结、化痰消肿。除此,壳亦可作肥料。海星及海燕等干制品可作肥料,并能入药,有清热解毒、平肝和胃、补肾滋阴的功能。自海星中提取的粗皂苷对大白鼠的实验性胃溃疡有较强的愈合作用;海星卵为研究受精及早期胚胎发育的好材料。蛇尾为一些冷水性底层鱼(鳕鱼)的天然饵料。
海胆喜食海藻,故为藻类养殖之害;有些种类的棘有毒,可造成对人类的危害。海星喜食双壳类,据记载,一个30天的小海星,6天中吃了50多个小海螂;一个成体海星一天吞食和破坏牡蛎可达20多个,故海星为贝类养殖之敌害。
第五节 棘皮动物的系统发展
棘皮动物体呈辐射对称,但它们的幼虫为两侧对称,因此辐射对称是次生形成的。一些已绝迹的化石种类中有的为两侧对称体形,如出现于寒武纪地层中的海林擒类(Cystidea)化石和海蕾类(Blastoidea)化石。因此有人认为棘皮动物的祖先为两侧对称体形的对称幼虫(dipleurula),具有3对体腔囊,与现在生存的棘皮动物幼虫形态类似。也有一些人主张五触手幼虫(pentactula)为棘皮动物的祖先。五触手幼虫也为两侧对称体形,具3对体胶囊和围绕口的5条中空触手;5条触手似总担,为体腔囊的延伸,是形成水管系统的基础。五触手幼虫由于进化为固着生活,其体形逐步转化为辐射对称。有一部分以后再营自由生活,但其体形仍保持着辐射对称。海百合纲为最古老的一类,出现于寒武纪,泥盆纪以后逐渐衰落。它们大多数营固着生活,其形态特征与海林擒纲和海蕾纲相似,且海百合类的幼虫与海林擒类近似,故海百合纲可能来源于海林擒纲。海星纲与蛇尾纲体形一致,均为五辐射对称,这二类的演化关系较为接近。而海胆纲与蛇尾纲的幼虫均为长腕幼虫,在结构上相似,二者关系较近。但海胆纲心形目动物,体近心脏形,肛门位体后端,体形属两侧对称。故海胆纲是介于蛇尾纲和两侧对称体形的海参纲之间的一个类群。海参纲体呈蠕虫状,两侧对称,口与肛门位体的前后两端,是棘皮动物中特殊的一类。其樽形幼虫与海百合纲的樽形幼虫很相似,故与海百合纲有着较近的类缘关系。海参只有一个生殖腺,这是较为原始的性状。故海参纲可能在演化过程中较早的分出的一支。
棘皮动物不同于大多无脊椎动物,而与脊索动物一样,同属后口动物。次生体腔由肠腔囊发育形成,中胚层产生内骨骼,这也是脊索动物的特征。海参纲的耳状幼体与半索动物肠鳃类的柱头虫幼虫(tornaria)在结构上非常相似,因此棘皮动物是无脊椎动物中与脊索动物最为相似的类群。
附:毛颚动物门
毛颚动物体形似箭,较小,体长多在40mm以下,半透明。海产,绝大多数营浮游生活(Snadella属为底栖),为海洋中浮游动物的组成部分,运动迅速,并可跳跃。
毛颚动物体分为头、躯干和尾三部分,各部在体内有隔膜分开。头端圆,腹侧为一纵裂的口,左右两侧具几丁质的刚毛,可帮助摄食,有颚的功能。以小鱼或小的甲壳类为食。躯干部有1~2对侧鳍,尾部具一三角形的尾鳍。肛门位躯干部末端腹面(图12-20)。
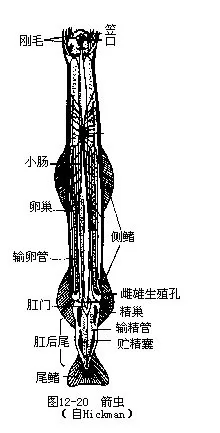
体腔为次生体腔,起源于体腔囊,被横隔分为头、躯干和尾三部分。体腔内充满体腔液,有运输功能。无循环系统和排泄系统。复层上皮,这与其他无脊椎动物完全不同。
毛颚类为雌雄同体,体后部两侧有卵巢一对,雌性生殖孔位体末端侧面;精巢一对,成熟精子由体壁破裂排出体外。多为异体受精,完全均等卵裂,经有腔囊胚,以内陷法形成原肠胚。于原肠胚前端形成口,故属后口动物。个体发育中尾部先分化,无幼虫期。
咽背侧有一脑神经节,以围咽神经连于体前腹侧的肠下神经节,形成一神经环。自脑神经节两侧向后伸出一对纵神经,至体中部与腹神经节相连。感官有眼一对,位头部。
毛颚动物约60多种,我国东海已记录21种,其中箭虫属(Sag-itta)有14种。肥胖箭虫(S.enflata)为东海的优势种,垂直分布为0m~250m,但99%个体在100m上层,其中75%在0m~50m,少数分布在250m~500m(林雅蓉,1985)。另外尚有龙翼箭虫(Pterosagitta draco),太平洋虫(Krohnitta pacifica)等;多分布在0m~50m上层。深水种有钩刺真虫(Eukrohnia hamata)。
毛颚动物的体腔形成及成体口的来源与后口动物相同,但其结构简单,复层上皮,体腔有隔膜等特点,可能为原始后口动物中极特化的一支,与其他后口动物的类缘关系不很密切。
复习题
1.棘皮动物门的主要特征是什么?
2.毛颚动物门的主要特征是什么?
3.为什么说棘皮动物、毛颚动物为无脊椎动物中的高等类群?
4.了解棘皮动物和毛颚动物的经济意义。
5.试述棘皮动物的系统发育及其对了解动物演化的意义。
本文标题:第十二章 棘皮动物门(Echinodermata) 附:毛颚动物门
手机页面:http://m.dljs.net/dlsk/dongwuxue/59421.html
本文地址:http://www.dljs.net/dlsk/dongwuxue/59421.html
