(一)脊索动物门的主要特征
脊索动物是动物界中最高等的一门,是与人类关系最密切的动物类群。现存种类不论在外部形态和内部结构上,或是生活方式方面,都存在着极其明显的差异,但作为同属一门的动物来说,几无例外地于其个体发育的某一时期或整个生活史中,都具有如下几点主要的共同特征。
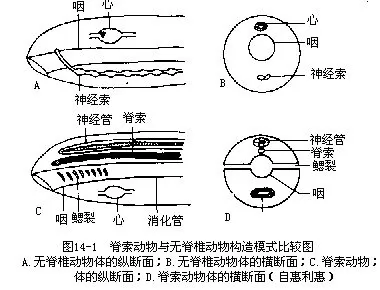
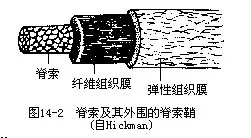
1.脊索(notochord)是背部起支持体轴作用的一条棒状结构,介于消化道和神经管之间。脊索来源于胚胎期的原肠背壁,经加厚、分化、外突,最后脱离原肠而成脊索。脊索由富含液泡的脊索细胞组成,外面围有脊索细胞所分泌而形成的结缔组织性质的脊索鞘(notochordal sheath)。脊索鞘常包括内外两层,分别为纤维组织鞘(fibrous sheath)和弹性组织鞘(elastic sheath)。充满液泡的脊索细胞由于产生膨压,使整条脊索既具弹性,又有硬度,从而起到骨骼的基本作用(图14-2)。低等脊索动物中,脊索终生存在或仅见于幼体时期。高等脊索动物只在胚胎期间出现脊索,发育完全时即被分节的骨质脊柱(vertebral column)所取代。组成脊索或脊柱等内骨骼(endoskeleton)的细胞,都能随同动物体发育而不断生长。而无脊椎动物则缺乏脊索或脊柱等内骨骼,通常仅身体表面被有几丁质等外骨骼(exoskeleton)。
2.背神经管(dorsal tubular nervecord)脊索动物神经系统的中枢部分是一条位于脊索背方的神经管,由胚体背中部的外胚层下陷卷褶所形成。背神经管在高等种类中前、后分化为脑和脊髓。神经管腔(neurocoele)在脑内形成脑室(cerebral ventricle),在脊髓中成为中央管(centralcanal)。无脊椎动物神经系统的中枢部分为一条实性的腹神经索(ventral nerve cord),位于消化道的腹面。
3.咽鳃裂(pharyngeal gill slits)低等脊索动物在消化道前端的咽部两侧有一系列左右成对排列、数目不等的裂孔,直接开口于体表或以一个共同的开口间接地与外界相通,这些裂孔就是咽鳃裂。低等水栖脊索动物的鳃裂终生存在并附生着布满血管的鳃,作为呼吸器官,陆栖高等脊索动物仅在胚胎期或幼体期(如两栖纲的蝌蚪)具有鳃裂,随同发育成长最终完全消失。无脊椎动物的鳃不位于咽部,用作呼吸的器官有软体动物的栉鳃以及节肢动物的肢鳃、尾鳃、气管等。
4.脊索动物的心脏及主动脉 位于消化道的腹面,循环系统为闭管式。无脊椎动物的心脏及主动脉在消化道的背面,循环系统大多为开管式。
5.极大多数脊索动物于肛门后方有肛后尾(post-anal tail)。无脊椎动物的肛孔常开口在躯干部的末端。
在上述特征中,具有脊索、背神经管和咽鳃裂是区别脊索动物和无脊椎动物最主要的3个基本特征。此外,脊索动物还有一些性状同样也见于高等无脊椎动物的,例如三胚层、后口、存在次级体腔、两侧对称的体制、身体和某些器官的分节现象等。这些共同点表明脊索动物是由无脊椎动物进化而来的。
(二)脊索的出现在动物演化史上的意义
脊索的出现是动物演化史中的重大事件,使动物体的支持、保护和运动的功能获得“质”的飞跃。这一先驱结构在脊椎动物达到更为完善的发展,从而成为在动物界中占统治地位的一个类群。
脊索(以及脊柱)构成支撑躯体的主梁,是体重的受力者,使内脏器官得到有力的支持和保护,运动肌肉获得坚强的支点,在运动时不致由于肌肉的收缩而使躯体缩短或变形,因而有可能向“大型化”发展。脊索的中轴支撑作用也使动物体更有效地完成定向运动,对于主动捕食及逃避敌害都更为准确、迅捷。脊椎动物头骨的形成、颌的出现以及椎管对中枢神经的保护,都是在此基础上进一步完善化的发展。
第二节 脊索动物分类概述
现存的脊索动物约有4万多种,分属于3个亚门,简述如下:
一、尾索动物亚门(Subphylum Urochordata)
脊索和背神经管仅存于幼体的尾部,成体退化或消失。体表被有被囊(tunic)。常见种类有各种海鞘和住囊虫,营自由生活或固着生活。有些种类有世代交替现象。本亚门包括尾海鞘纲(Appendiculariae)、海鞘纲(Ascidiacea)、樽海鞘纲(Thaliacea)等。
二、头索动物亚门(Subphylum Cephalochordata)
脊索和神经管纵贯于全身的背部,并终生保留。咽鳃裂众多。本亚门仅头索纲(Cephalo-chorda)一个类群,体呈鱼形,体节分明,表皮只有一层细胞,头部不显,故称无头类(Acrania)。
三、脊椎动物亚门(Subphylum Vertebrata)
脊索只在胚胎发育阶段出现,随后或多或少地被脊柱所代替。脑和各种感觉器官在前端集中,形成明显的头部,故称有头类(Craniata)。本亚门包括:
(一)圆口纲(Cyclostomata) 无颌,缺乏成对的附肢,单鼻孔,脊索及雏形的椎骨(verte-brae)并存。又名无颌类(Agnatha)。
(二)鱼纲(Pisces) 出现上、下颌,体表大多被鳞,鳃呼吸,成对的前后肢形成适于水生生活的胸鳍和腹鳍。本纲与更高等的四足类(Tetrapoda)脊椎动物合称为有颌类(Gnathostomata)。
(三)两栖纲(Amphibia) 皮肤裸露,幼体用鳃呼吸,以鳍游泳,经过变态后的动物上陆生活,营肺呼吸和以五趾型附肢(pentadactyl limb)运动。
(四)爬行纲(Reptilia) 皮肤干燥,外被角质鳞、角盾或骨板。心脏有二心房、一心室或近于两心室。本纲与鸟纲、哺乳纲在胚体发育过程中出现羊膜(amnion),因而合称为羊膜动物(Amniota),其他各纲脊椎动物则合称为无羊膜动物(Anamniota)。
(五)鸟纲(Aves) 体表被羽(feather),前肢特化成翼,恒温,卵生。
(六)哺乳纲(Mammalia) 身体被毛,恒温,胎生,哺乳。
第三节 尾索动物亚门
尾索动物和头索动物两个亚门是脊索动物中最低级的类群,总称为原索动物(Protochorda-ta)。身体包在胶质(gelatinous)或近似植物纤维素成分的被囊中,至少在幼体时期尾部具有脊索及神经管,所以称为尾索动物或被囊动物(tunicate)。全世界约有1370多种,常见种类有柄海鞘(Styela clava)、海樽(Ascida)、玻璃海鞘(Ciona)、菊花海鞘(Botryllus)等,分布遍及世界各地的海洋。体呈袋形或桶状,包括单体或群体两个类型,绝大多数无尾种类只在幼体时期自由生活,成体于浅海潮间带营底栖固着生活,少数终生有尾种类在洋面上营漂浮式的自由游泳生活。体表有入水孔(incurrent siphon)和出水孔(excurrent siphon),咽壁有数量不等的鳃裂,咽外围有宽大的围鳃腔(peribranchial cavity),与出水管孔相通。一般为雌雄同体(hermaphroditism),异体受精。由于卵和精子并不同时成熟,所以避免了自体受精。营有性生殖,也营无性的出芽生殖,樽海鞘的生活史中甚至还有复杂的世代交替现象。除个别种类外,受精卵都先发育成善于游泳的蝌蚪状幼体,再行变态发育。
早在2千多年前,尾索动物就已经被记载和描述,曾先后隶属于无脊椎动物中的蠕虫类、拟软体动物、苔藓动物或软体动物,直到1866年俄国学者柯瓦列夫斯基仔细地研究了海鞘的胚胎发育及其变态后,才正式判定其正确的分类地位,置于脊索动物门。
一、代表动物——柄海鞘(Styela clava)
海鞘是尾索动物亚门中最主要的类群,约占全部种数的90%以上。柄海鞘是海鞘类中的优势种,经常与盘管虫(Hydroides)、藤壶(Balanus)及苔藓虫(Bugula)等附着在一起,固着在码头、船坞、船体,以及海水养殖的海带筏和扇贝笼上,被作为沿海污损生物的重要指标种。柄海鞘的身体构造及变态过程在本月动物中具有一定的代表性,现以此为例简述其主要特征。
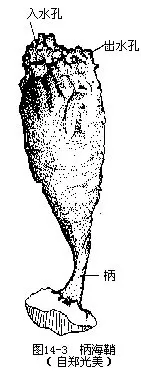
(一)外形和生活方式 柄海鞘的成体呈长椭圆形,基部以柄附生在海底或被海水淹没的物体上,另一端有2个相距不远的孔:顶端的一个是入水孔,孔内通消化管而中间有一片筛状的缘膜,其作用是滤去粗大的物体,只容许水流和微小食物进入消化道;位置略低的一个是出水孔(图14-3)。从胚胎发生和幼体变态的过程来看,两孔之间是柄海鞘的背部,对应的一侧为腹部。一般情况下,水流从入水孔进入而由出水孔排出,当受惊扰或刺激时,则可引起体壁骤然收缩,体内积贮的水分别从2个孔中似乳汁般同时喷射而出,故在山东省沿海一带俗称海奶子;刺激缓解后,身体又逐渐恢复原状。它们除了可以成簇密集生活外,还能附着在同种的其它个体上,同时本身又可以被别的个体所附着,呈现垒叠的聚生现象。
(二)内部构造
1.体壁 柄海鞘的体壁即是包藏内部器官的外套膜(mantle),外套膜除了表面一层外胚层的上皮细胞外,还参杂着来源于中胚层的肌肉纤维,以支配身体及出、入水孔的伸缩和开关。体壁能分泌一种化学成分类似植物纤维素的被囊素(tunicin),并由此形成包围在动物体外的被囊,这就是被囊动物名称的由来。在整个动物界中,体壁能分泌被囊素的动物,至今仅发现于尾索动物和少数原生动物。外套膜在入水孔和出水孔的边缘处与被囊汇合,汇合处有环形括约肌控制管孔的启闭。内部器官中只有咽的上缘及腹面的一部分与外套膜愈合。柄海鞘的被囊表面通常不易被其它动物所附着,但是同种个体却能重叠附生,这对种群的繁衍显然是有积极意义的(图14-4)。
2.消化和呼吸系统 入水孔的底部有口,通过四周长有触手的缘膜就是宽大的咽,咽几乎占据了身体的大半部(3/4),咽壁被许多细小的鳃裂所贯穿。从口进入咽内的水流经过鳃裂,到达围着咽外的围鳃腔中,然后经出水孔排出。围鳃腔是由身体表面陷入内部所形成的空腔,因其不断扩大,从而将身体前部原有的体腔逐渐挤小,最终在咽部完全消失。由于鳃裂的间隔里分布着丰富的毛细血管,因此当水流携带着食物微粒通过鳃裂时就能进行气体交换,完成呼吸作用。咽腔的内壁生有纤毛,其背、腹侧的中央各有一沟状结构,分别称为背板(dorsal lamina或咽上沟epipharyngeal groove)和内柱(endostyle),沟内有腺细胞和纤毛细胞;背板和内柱上下相对,在咽的前端以围咽沟(peripharyngeal groove)相连,腺细胞能分泌粘液,使沉入内柱的食物粘聚成团,由沟内的纤毛摆动,将食物团从内柱推向前行,经围咽沟沿背板往后导入食道、胃及肠进行消化。肠开口于围鳃腔,不能消化的残渣通过围鳃腔,随水流经出水孔排出体外。
3.循环方式和排泄器官心脏位于身体腹面靠近胃部的围心腔(pericardial cavity)内,藉围心膜的伸缩而搏动。心脏两端各发出一条血管,前端一条为鳃血管,沿咽腹发出分支到鳃裂间的咽壁上;后端一条称肠血管,分支到各内脏器官并注血进入器官组织的血窦之间,所以是开管式的血液循环。柄海鞘具有特殊的可逆式血液循环流向,即心脏收缩有周期性间歇,当它的前端连续搏动时,血液不断地由鳃血管压出至鳃部,接着心脏有短暂的停歇,容纳鳃部的血液流回心脏,然后于其后端开始搏动,将血液注入肠血管而分布到内脏器官的组织间。因此,柄海鞘的血管既无动脉和静脉之分,血液也无固定的单向流动方向,这种独特的血液循环方式在动物界中是绝无仅有的。
柄海鞘无专门的排泄器官,仅在肠附近有一堆具排泄机能的细胞,称为小肾囊(renal vesicles),其中常含尿酸结晶。
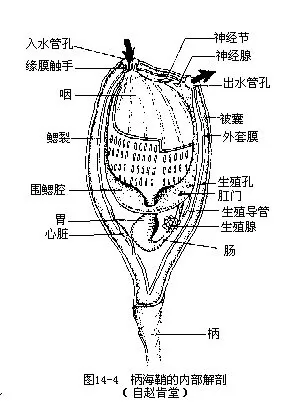
4.神经柄海鞘的成体营固着生活,神经系统和感觉器官均甚退化,中枢神经只是一个没有内腔的神经节(nervus ganglion),圆而坚硬,状如小瘤,位于入水孔和出水孔之间的外套膜壁内,由此分出若干神经分支到身体各部,神经节旁有一无色透明而略为膨大的神经腺(neural gland,相当高等动物的脑下腺hypophy-sis)。无专门的感觉器官,仅于入水管孔、出水管孔的缘膜和外套膜上有少量散在的感觉细胞。
5.生殖系统雌雄同体,生殖腺位于肠环间和外套膜内壁上。精巢大,呈分支状,为乳白色颗粒状小块;卵巢长管状,呈淡黄色,内含许多圆形的卵细胞;两者紧贴重叠,分别以单根生殖导管(gonoduct)将成熟的性细胞输入围鳃腔,然后经出水管孔排至体外,或在围鳃腔内与另一海鞘的生殖细胞相遇受精。
(三)幼体及变态柄海鞘成体的形态结构与典型的脊索动物有很大差异。然而,它的幼体外形酷似蝌蚪并具有脊索动物3个主要特征。幼体长约0.5mm,尾内有发达的脊索,脊索背方有中空的背神经管,神经管的前端甚至还膨大成脑泡(cerebral vesicle),内含眼点和平衡器官等;消化道前段分化成咽,有少量成对的鳃裂;身体腹侧有心脏。
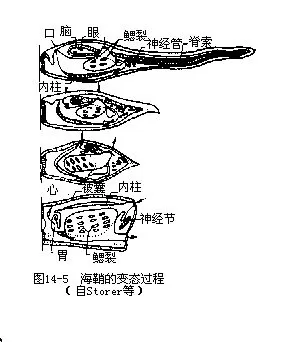
幼体经过几小时的自由生活后,就用身体前端的附着突起(adhesivepapillae)粘着在其它物体上,开始其变态。在变态过程中,海鞘幼体的尾连同内部的脊索和尾肌逐渐萎缩,并被吸收而消失,神经管及感觉器官也退化而残存为一个神经节。与此相反,咽部却大为扩张,鳃裂数急剧增多,同时形成围绕咽部的围鳃腔;附着突起也为海鞘的柄所替代。附着突起背面因生长迅速,把口孔的位置推移到另一端(背部),于是造成内部器官的位置也随之转动了90°~180°的角度。最后,由体壁分泌被囊素构成保护身体的被囊,使它从自由生活的幼体变为营固着生活的柄海鞘(图14-5)。柄海鞘经过变态,失去了一些重要的构造,形体变得更为简单,这种变态称为逆行变态(retrogressive metamorphosis)。
二、尾索动物分类
本亚门是脊索动物中最低等的类群,遍布世界各个海洋,约1370多种,分属于3纲(图14-6),我国已知有14种左右。
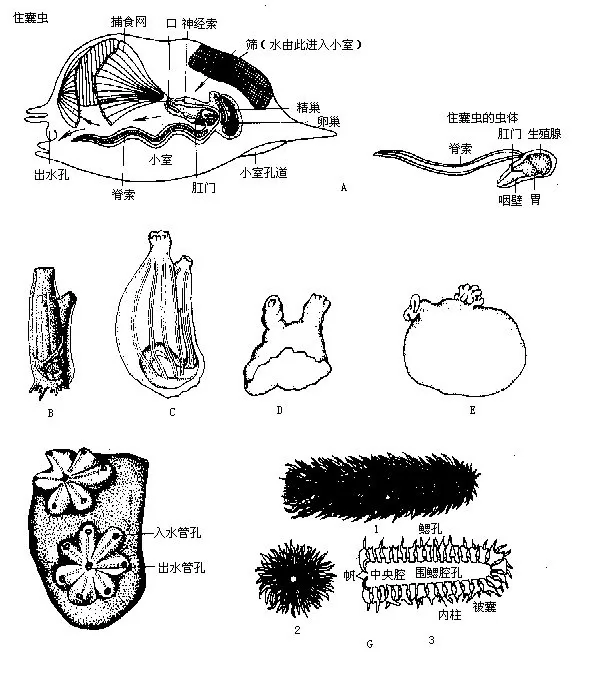
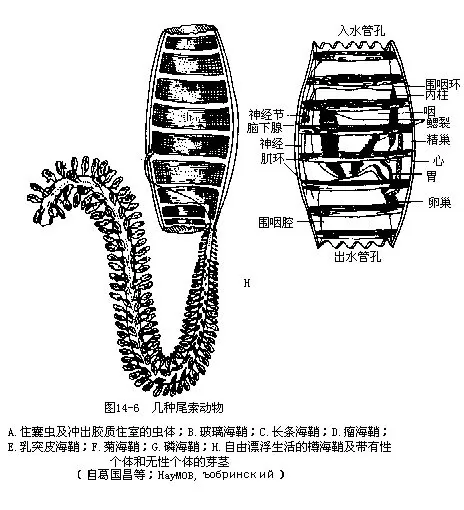
(一)尾海鞘纲(Appendiculariae)本纲是尾索动物中的原始类型,共1目3科60余种。体长数毫米至20mm,代表动物为住囊虫(Oikopleura)和巨尾虫(Megalocercus huxleyi)等。尾海鞘纲与本亚门中其他2纲的主要区别是:体外无被囊,只有两个直接开口体外的鳃裂而缺乏国鳃腔,终生保持着带有长尾的幼体状态(neotonous),大多在沿岸浅海中营自由游泳生活。生长发育过程中无逆行变态,故又名幼形纲(Larva-cea)。住囊虫包藏在由皮肤分泌的胶质住囊(gelatinous‘house’)内,住囊有入水孔和出水孔,住囊虫在囊中借助内有脊索和神经索的尾巴摆动进水,并使囊中的水由出水孔排出,推动动物体前进,同时通过虫体口外特有的网筛(filter),从流水中滤取微小的浮游生物作为食物。每隔数小时,住囊的出、入水孔就将被堵塞。此时住囊虫即激烈挥动长尾,从特殊的应急出口(或称小室孔道 escape hatch)破囊冲出至海中,并在很短的时间里再形成新的住囊。
我国至今尚未发现本纲动物。
(二)海鞘纲(Ascidiacea)种类繁多,约有1250种,包括单体和群体2种类型,附着于水下物体或营水底固定生活。单体型种类的最大体长可达200余mm,群体的全长可超过0.5m以上。群体型种类的许多个体都以柄相连,并被包围在一个共同的被囊内,但分别以各自的入水孔进水,有共同的排水口,如群体海鞘(Diplosoma)。
广布于我国的海鞘纲动物有米氏小叶鞘(Leptoclinum mitsukurii)、星座美洲海鞘(Amarou-cium constellatum)、长纹海鞘(Ascidia longistriata)、玻璃海鞘(Ciona intestinalis)、3种菊海鞘(Botryllus ssp.)、瘤海鞘(Styela canops)、乳突皮海鞘(Molgula manhattensis)、龟甲海鞘(Chely-osoma)、西门登拟菊海鞘(Botrylloides simodensis)等。乳突皮海鞘和柄海鞘是连云港以北黄、渤海沿岸污损生物中的优势种,瘤海鞘大量出现在香港、西沙永兴岛和琛航岛等南海海域内。
(三)樽海鞘纲(Thaliacea)本纲动物大多是营自由游泳生活的漂浮型海鞘,体呈桶形或樽形,咽壁有2个或更多的鳃裂。成体无尾,入水孔和出水孔分别位于身体的前后端。被囊薄而透明,囊外有环状排列的肌肉带,肌肉带自前往后依次收缩时,流进入水孔的水流即可从体内通过出水孔排出,以此推动樽海鞘前进,并在此过程中完成摄食和呼吸作用。生活史较复杂,繁殖方式是有性与无性的世代交替。樽海鞘纲约有65种,代表动物有樽海鞘(Doliolum deuticula-tum),我国厦门沿海曾发现过小海樽(Dolioletta natilnalis)。磷海鞘(Pyrosoma atlanticum)为群体型种类,身体总长度可达200mm~600mm,因其口孔内缘有磷光器,漂浮游泳时能发出闪烁的磷光而得名,别名火体虫。
三、尾索动物的演化
尾索动物是最低等的脊索动物,与高等脊索动物存在着演化上的亲缘关系,两者可能都是从类似海鞘幼虫型营自由生活的共同祖先——原始无头类动物演化而来。这类原始无头类动物不但将幼体时期的尾和自由游泳的生活方式保留到成体,甚至还消失了生活史中营固着生活的阶段,并通过幼态滞留及幼体性成熟途径发展为头索动物和脊椎动物。尾索动物是在进化过程中适应特殊生活方式的一个退化分支,除保留滤食的咽及营呼吸作用的咽鳃裂外,大多数种类已在变态中失去所有的进步特征,并向固着生活的方向发展。
第四节 头索动物亚门
头索动物是一类终生具有发达脊索、背神经管和咽鳃裂等特征的无头鱼形脊索动物。包括头索纲(Cephalochorda,又名狭心纲Leptocardii)、鳃口科(Branchiostomidae),约30种,分隶于文昌鱼和偏文昌鱼(Asymmetron)2个属。头索动物分布很广,遍及热带和温带的浅海海域,其中尤以北纬48°至南纬40°之间的沿海地区数量较多。
头索动物的脊索不但终生保留,且延伸至背神经管的前方,故称头索动物。又因本亚门动物都缺乏真正的头和脑,所以又称无头类。我国厦门、青岛等地所产的文昌鱼( Branchiostomabelcheri),可作为头索动物的代表。
一、代表动物——白氏文昌鱼( Branchiostoma belcheri)
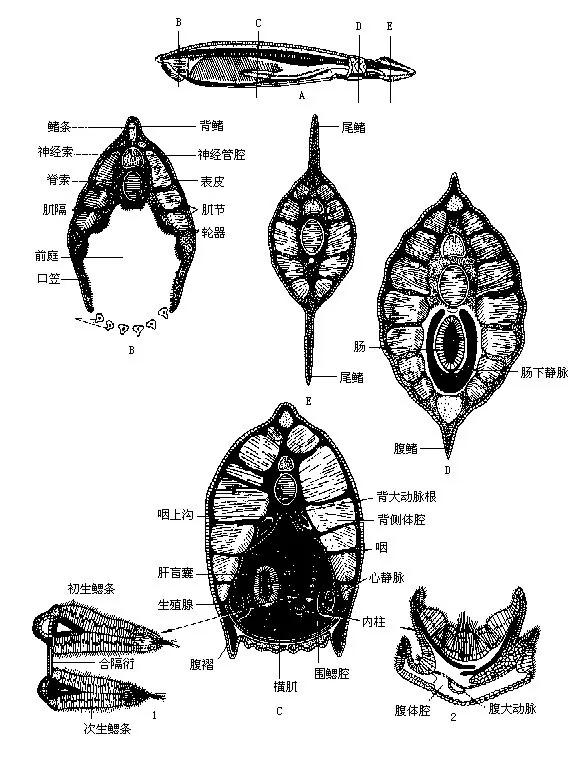
(一)外形和生活方式文昌鱼的体形略似小鱼,无明显的头部,左右侧扁,半透明,可隐约见到皮下的肌节(myomere)和腹侧块状的生殖腺;身体两端尖出,故有双尖鱼(Amphioxus)之称,又因其尾形很像矛头而名海矛。一般体长约50mm,但产于美国的加州文昌鱼(Branchiostomacaliforniense)可超过100mm,是该属中已知个体最大的一种。前端的腹面为一漏斗状的口笠(oral hood),口笠内为前庭(vestibule),内壁有轮器(wheel organ),由前庭引向位于一环形缘膜(velum)中央的口。口笠和缘膜的周围分别环生触须(cirri)及缘膜触手(velar tentacle),具有保护和过滤作用,可阻挡粗砂等物随水流进入口中。整个背面沿中线有一条低矮的背鳍(dorsalfin),往后与高而绕尾的尾鳍(caudal fin)相连。此外在肛门之前还有肛前鳍(preanal fin)。无偶鳍,只在身体前部的腹面两侧各有一条由皮肤下垂形成的纵褶,称为腹褶(metapleura fold)。腹褶和肛前鳍的交界处有一腹孔(atripore),是咽鳃裂排水的总出口,故又名围鳃腔孔(图14-7)。
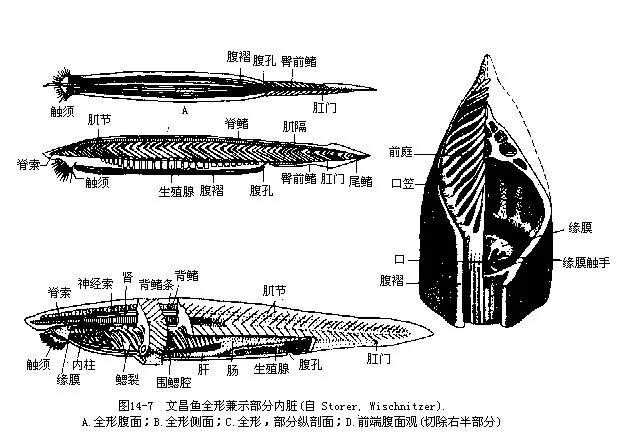
文昌鱼喜栖浅海水质清澈的沙滩上,平时很少活动,常把身体半埋于沙中,前端露出沙外,或者左侧贴卧沙面,借水流携带矽藻等浮游生物进入口内。夜间较为活跃,凭藉体侧肌节的交错收缩,在海水中作短暂的游泳。寿命约2年8个月左右。5~7月为生殖季节,一生中可繁殖3次,其中以最后一次产卵最多。福建沿海海区曾是我国文昌鱼生息繁衍的理想场所。1932—1956年,文昌鱼在同安县一地的捕捞区面积约有22km2,每升沙样中文昌鱼的数量达357条,当时因年产量约57吨而高居世界首位。1956年,由于厦门高(崎)集(美)海堤建成后,海况(切断了环岛的海流、沙地面积大幅度减少、盐度降低、水流减缓、淤泥沉积等)发生很大变化,再加上围海造田和乱捕滥捞等原因,严重地破坏了文昌鱼的生态环境,因而造成数量急剧下降的恶果,年产量减少了40%。1970年,当地的收购量甚至降低到只有1t左右。1986年,文昌鱼在这一海域的分布区已缩小到0.5km2,而每L沙样中的数量进一步减少到3条,使原来名闻遐迩的文昌鱼产地,在20世纪半个世纪内变得完全失去了水产捕捞价值。
(二)内部构造特征
1.皮肤 皮肤薄而半透明,由单层柱形细胞的表皮和冻胶状结缔组织的真皮两部分构成,表皮外覆有一层角皮层(cuticle)。表皮外在幼体期生有纤毛,成长后则消失殆尽。
2.骨骼 文昌鱼尚未形成骨质的骨骼,主要是以纵贯全身的脊索作为支持动物体的中轴支架。脊索外围有脊索鞘膜,并与背神经管的外膜、肌节之间的肌隔、皮下结缔组织等连续。脊索细胞呈扁盘状,其超显微结构与双壳类软体动物的肌细胞比较相似,收缩时可增加脊索的硬度。此外,在口笠触须、缘膜触手、轮器内部也都有角质物支持,奇鳍和鳃裂的鳍条(fin rays)及鳃条(gill bar)由结缔组织支持。
3.肌肉 文昌鱼背部的肌肉厚实而腹部比较单薄,与无脊椎动物周身体壁厚薄均匀的情况不同。全身主要的肌肉是60多对按节排列于体侧的“V”字形肌节(myomere),尖端朝前,肌节间被结缔组织的肌隔(myocomma)所分开。两侧的肌节互不对称,便于文昌鱼在水平方向作弯曲运动。此外,还有分布在围鳃腔腹面的横肌和口缘膜上的括约肌等,控制围鳃腔的排水及口孔的大小。
4.消化和呼吸器官 文昌鱼靠轮器和咽部纤毛的摆动,使带有食物微粒的水流经口入咽,食物被滤下留在咽内,而水则通过咽壁的鳃裂至围鳃腔,然后由腹孔排出体外。作为收集食物和呼吸场所的咽部极度扩大,几乎占据身体全长的1/2,咽腔内的构造与尾索动物相似,也具有内柱、咽上沟和围咽沟等。文昌鱼幼体的鳃裂直接开口于体表,后来形成围鳃腔,以腹孔作为咽部鳃裂的总出水口。
咽内的食物微粒被内柱细胞的分泌物粘结成团,再由纤毛运动使它从后向前流动,经围咽沟转到咽上沟,往后推送进入肠内。肠为一直管,向前伸出一个盲囊,突入咽的右侧,称为肝盲囊(hepatic diverticulum),能分泌消化液,与脊椎动物的肝脏为同源器官。食物团中的小微粒可进入肝盲囊,被肝盲囊细胞所吞噬,营细胞内消化,大微粒在肠内分解成小微粒后,也转到肝盲囊中进行细胞内消化,未消化的物质由肝盲囊重返肠中,在后肠部进行消化和吸收。肠的末端开口于身体左侧的肛门。
咽腔是文昌鱼完成呼吸作用的部位。咽壁两侧有60多对鳃裂,彼此以鳃条分开,鳃裂内壁布有纤毛上皮细胞和血管。水流进入口和咽时,藉纤毛上皮细胞的纤毛运动,通过鳃裂,并使之与血管内的血液进行气体交换,最后,水再由围鳃腔经腹孔排出体外。有人认为文昌鱼纤薄的皮肤也具有直接从水中摄取氧气的能力。
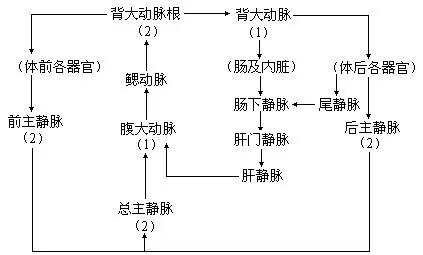
5.血液循环文昌鱼循环系统属于闭管式,即血液完全在血管内流动,这种情形与脊椎动物基本相同。无心脏,但是具搏动能力的腹大动脉(ventral aorta),因而被称为狭心动物。由腹大动脉往两侧分出许多成对的鳃动脉(branchial arteries)进入鳃隔,鳃动脉不再分为毛细血管,它在完成气体交换的呼吸作用后,于鳃裂背部汇入2条背大动脉根。背大动脉根内含多氧血,往前流向身体前端,向后则由左、右背大动脉根合成背大动脉(dorsal aorta),再由此分出血管到身体各部。血液无色,也没有血细胞和呼吸色素,动脉中的血液通过组织间隙进入静脉。从身体前端返回的血液通过体壁静脉(parietal vein)注入一对前主静脉(anterior cardinal vein);尾的腹面有一条尾静脉(caudal vein),收集一部分身体后部回来的血液,进入肠下静脉(subintestinal vein),大部分血液则流进2条后主静脉(posterior cardinal vein)。左、右前主静脉和两条后主静脉的血液全部汇流至一对横形的总主静脉(common cardinal vein),或称居维叶氏管(ducts Cuvieri)。左、右总主静脉会合处为静脉窦(sinus venosus),然后通入腹大动脉。从肠壁返回的血液由毛细血管网集合成肠下静脉,尾静脉的部分血液也注入其中;肠下静脉前行至肝盲囊处血管又形成毛细管网,由于这条静脉的两端在肝盲囊区都形成毛细血管,因此称作肝门静脉(hepatic portal vein)。由肝门静脉的毛细血管再一次合成肝静脉(hepatic vein)并将血液汇入静脉窦内。
文昌鱼的血液循环路线,可以表示如下:
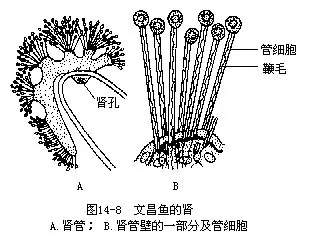
6.排泄器官排泄器官由数10对按节排列的肾管(nephridium)组成,位于咽壁背方的两侧,其结构和机能与扁形动物及环节动物的原肾比较近似。每个肾管是一短而弯曲的小管,弯曲的腹侧有单个开口于围鳃腔的肾孔(nephrostome),弯管的背侧连接着5~6束与肾管相通的管细胞(solenocytes)。管细胞由体腔上皮细胞特化而成,其远端呈盲端膨大,紧贴体腔,内有一长鞭毛。代谢废物通过体腔液渗透进入管细胞,经鞭毛的摆动到达肾管,再由肾孔送至围鳃腔,随水流排出体外(图14—8)。此外,在咽部后端背部的左右,各有一个称为褐色漏斗(brown funnel)的盲囊,有人认为有排泄功能,但也有人推测可能是一种感受器。
7.神经系统文昌鱼的中枢神经是一条纵行于脊索背面的背神经管,神经管的前端内腔略为膨大,称为脑泡(cerebralvesicle)。幼体的脑泡顶部有神经孔与外界相通,长成后完全封闭。神经管的背面并未完全愈合,尚留有一条裂隙,称为背裂(dorsal fissure)。周围神经包括由脑泡发出的2对“脑”神经和自神经管两侧发出的成对脊神经。神经管在与每个肌节相应的部位,分别由背、腹发出一对背神经根及几条腹神经根,或简称背根(dorsal root)和腹根(ventral root)。背根和腹根在身体两侧的排列形式与肌节一致,左右交错而互不对称,且其背根和腹根之间也不像脊椎动物那样联结成一条脊神经。背根是兼有感觉和运动机能的混合性神经,接受皮肤感觉和支配肠壁肌肉运动;腹根专管运动,分布在肌肉上。
与文昌鱼少动生活方式相联系的是感觉器官很不发达,许多位于神经管两侧的黑色小点是文昌鱼的光线感受器,称为脑眼(ocelli)。每个脑眼由一个感光细胞和一个色素细胞构成,可通过半透明的体壁,起到感光作用。神经管的前端有单个大于脑眼的色素点(pigment spot),色素点又叫眼点(eye spot),但无视觉作用,有人认为是退化的平衡器官,有人则以为有遮挡阳光使脑眼免受阳光直射的作用。此外,全身皮肤中还散布着零星的感觉细胞,其中尤以口笠、触须和缘膜触手等处较多。
8.生殖系统雌雄异体。生殖腺附生于围鳃腔两侧的内壁上,是26对左右厚壁的矩形小囊,性成熟时可根据文昌鱼的精巢为白色或卵巢呈现淡黄色进行雌雄鉴别。成熟的精子和卵都是通过生殖腺壁的破口释出,坠入围鳃腔,再随同水流由腹孔排出,在海水中完成受精作用(图14-9)
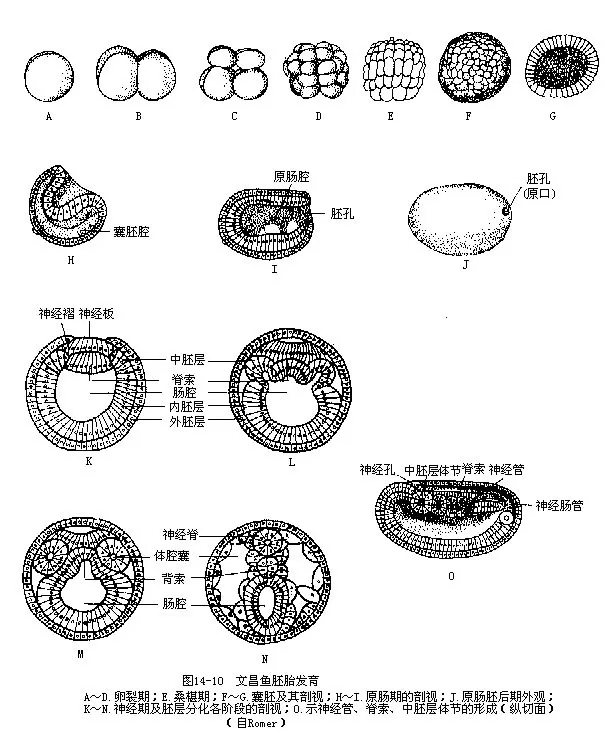
(三)胚胎发育和变态文昌鱼在6~7月产卵,通常产卵和受精都在傍晚进行。卵小而含卵黄少,为均黄卵(isolecithal egg),卵径0.1mm~0.2mm。文昌鱼的发育需经历受精卵—桑椹胚—囊胚—原肠胚—神经胚各个时期,才孵化成幼体。
受精卵进行几乎均等的全分裂(holoblastic),经过多次细胞分裂后,使许多细胞结成一个形似实心圆球的桑椹胚(morula)。桑椹胚在继续细胞分裂的同时,中心的细胞逐渐向胚体表面迁移,从而变成一个内部充满胶状液的空心囊胚(blastopore),囊胚中的腔为囊胚腔(blastocoel),囊胚上端的细胞略小,称动物极(animal pole),细胞较大的下端是植物极(vegetative pole)。接着,囊胚的植物极大细胞向内陷入以至与上端动物极细胞的内壁互相紧贴,囊胚腔因受挤压而消失,被新出现的原肠腔(archenteron)所代替。原肠腔以植物极细胞内陷处的胚孔(或称原口 blas-topore)与外界相通,这里相当于胚体的后端,相对的另一端为前端。此时胚胎已形成内、外两层细胞,分别称为内胚层(endoderm)和外胚层(ectoderm),胚体表面长有纤毛并能在胚膜中进行回旋运动。胚胎发育到这个阶段称为原肠胚(gastrula)。
原肠胚的后期开始胚体延长、产生中枢神经、胚层进一步分化及各器官系统的形成。
原肠胚自前端沿背中线至胚孔的外胚层下陷成神经板(neural plate),与此相邻的两侧外胚层同神经板脱离,互相靠拢而完全愈合,是将来的表皮部。下陷到表皮内的神经板首先在板的两侧往上隆起成神经褶(neural fold),然后卷合围成背面留有一条纵裂的神经管(neural tube),管内为神经管腔(neurocoel)。其前端以神经孔(neuropore)和外界相通,后端经胚孔与原肠相通成神经肠管(neurenteric canal)。成体时,神经孔关闭成嗅窝,而神经肠管也闭塞不通并在胚孔部形成肛门,此时的胚胎称为神经胚(neurula)。
在背神经管形成的同时,脊索和中胚层也在形成。原肠背面正中出现一条纵行的隆起实体,即脊索中胚层,它与原肠分离后发育成脊索。脊索两侧各有一列按节排列和彼此连接的体腔囊(coelomic sac),这就是新发生的中胚层,体腔囊中的每个空腔即体腔(coelom)。文昌鱼身体前部的中胚层是以体腔囊的方式所形成,与棘皮动物及半索动物相同,身体后部中胚层的发生方式又与脊椎动物一致,由此可见文昌鱼在两大类动物中处于过渡性质的居中地位,对动物学的一些基本理论研究方面,可以提供许多有价值的材料。
随着每个体腔囊的发育,又分化成背、腹两部分。背部称体节(somite),腹部称侧板(lateralplate),体节内的体腔以后自行消失,而侧板内的体腔最初因体腔囊分节彼此独立存在,后来由于体腔囊壁前后沟通,才在体内形成一个完整的体腔。这种体腔叫作次级体腔,是真正由中胚层所构成的体腔。
体节的内侧部分,又分化为生骨节(sclerotome),将来形成脊索鞘、背神经管外的结缔组织和肌隔等;体节的中部形成肌节;体节的外侧部分以后形成皮肤的真皮。侧板的外层为体壁中胚层(somatic mesoderm),将来发育成紧贴着体腔壁的腹膜或体腔膜(peritoneum);内层称脏壁中胚层(splanchnic mesoderm),以后形成肠管外围的组织。脏壁中胚层在肠管前段的背侧发生出分节排列的指状突起,即未来的肾管。体壁与侧板交界处的体腔壁上也发生突起,以后自此发育出文昌鱼的生殖腺(图14—10)。
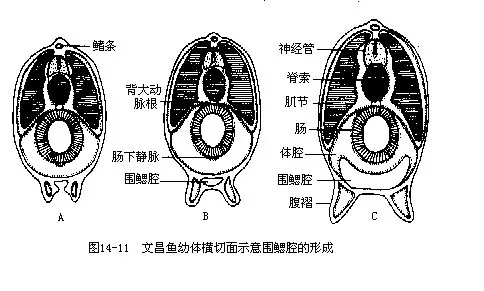
受精卵经过20多个小时后,文昌鱼的胚胎发育基本结束,全身披有纤毛的幼体就能突破卵膜,到海水中活动,此时有白天游至海底夜间升上海面进行垂直洄游的生活规律。幼体期约3个月,然后沉落海底进行变态。幼体在生长发育和变态的过程中,身体日益长大,出现前庭,鳃裂的数目因发生次生鳃条而增加了一倍,并由原来直接开口体外而变为通入新形成的围鳃腔中(图14—11)。一龄的文昌鱼体长约40mm,性腺发育成熟,可参与当年的繁殖。
二、头索动物在动物进化中的意义
头索动物的身体构造虽然比较简单,但已充分显示出是典型脊索动物的简化缩影。A.H.谢维尔曹夫根据文昌鱼的胚胎发育进行推测,头索动物的祖先似乎是一类身体非左右对称、无围鳃腔、鳃裂较少而直通体外、营自由游泳生活的动物。这样的动物称为原始无头类,很可能是头索动物和脊椎动物的共同始祖,它们在进化中由于适应不同的生活而分成两支演变,一支改进和发展了适于自由游泳生活的体制结构,演变成原始有头类,导向脊椎动物的进化之路;另一支往少动和底栖钻沙的生活方式发展,特化为旁支,演变成头索动物的鳃口科动物。曾有人先后在非洲、澳大利亚、斯里兰卡、苏门答腊的深海捕到过偏文昌鱼,从它所具有的无围鳃腔和肝盲囊、腹褶及鳃裂均不对称、只在身体右侧有生殖腺、口和肛门的位置偏左等特征看来,谢维尔曹夫有关文昌鱼演化的推论基本上是正确的。我国广东和海南等省的沿海也发现了短刀偏文昌鱼(Asymmetron cultellum)。
第五节 脊椎动物亚门(Vertebrata)
脊椎动物是脊索动物门中数量最多、结构最复杂、进化地位最高的一大类群,因而也是动物界中最进步的类群。这一群动物不但对于人类的物质生活、文化生活等有直接的关系,而且对于人类了解其自身也能提供许多宝贵的科学资料,因为人类在自然界的位置正是在脊椎动物之中。
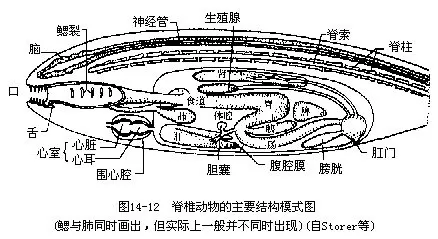
脊椎动物虽只是一个亚门,但因各自所处的环境不同,生活方式就显出千差万别,形态结构也彼此悬殊。然而高度的多样化并不能掩盖它们都属于脊索动物的共性,即在胚胎发育的早期都要出现脊索、背神经管和咽鳃裂。有些种类的幼体用鳃呼吸;有些种类即使是成体,也终生用鳃呼吸。脊椎动物的主要特征如图14—12。
1.出现了明显的头部神经管的前端分化成脑和眼、耳、鼻等重要的感觉器官,后端分化成脊髓。这就大大加强了动物个体对外界刺激的感应能力。由于头部的出现,脊椎动物又有“有头类”之称。
2.在绝大多数的种类中,脊索只见于发育的早期,以后即为脊柱(vertebralcolumn)所代替脊柱由单个的脊椎(vertebra)连接组成。脊椎动物就是因为具有脊椎而得名。脊柱保护着脊髓,其前端发展出头骨保护着脑。脊柱和头骨是脊椎动物特有的内骨骼的重要组成部分,它们和其他的骨骼成分一起,共同构成骨骼系统以支持身体和保护体内的器官。
3.原生的水生种类(即在系统发展上最多只达到鱼类阶段的动物)用鳃呼吸,次生的水生种类(即在系统发展上已超过鱼类阶段,因适应环境的关系又重新过水中生活,例如鲸类)及陆生种类只在胚胎期间出现鳃裂,成体则用肺呼吸。
4.除了圆口类之外,都具备了上、下颌(jaw)颌的作用在于支持口部,加强动物主动摄食和消化的能力。以下颌上举使口闭合的方式,为脊椎动物所特有,不见于其他类群。
5.完善的循环系统。出现了能收缩的心脏,促进血液循环,有利于生理机能的提高。在高等的种类(鸟类和哺乳类)中,心脏中的多氧血与缺氧血已完全分开,机体因得到多氧血的供应,所以能保持旺盛的代谢活动,使体温恒定,形成脊椎动物中所特有的恒温动物(温血动物)。
6.用构造复杂的肾脏代替了简单的肾管,提高了排泄系统的机能,使新陈代谢所产生的大量废物更有效地排出体外。
7.除了圆口类之外,都用成对的附肢(paired appendages)作为运动器官这就是水生种类的鳍(fin)和陆生种类的肢(limb)。这种成对的附肢,在整个脊椎动物中,数量不超过两对。所以,作为成对的鳍,只有胸鳍和腹鳍;作为成对的肢,也只有前肢和后肢。有少数种类失去了一对附肢(如河鲀没有腹鳍,鳗螈没有后肢)或甚至两对附肢都全部失去(如黄鳝、蛇)。这是一种次生现象,因为在它们的身上还不同程度地留有附肢的痕迹,说明它们是从有成对附肢的祖先演变而来。
复习题
1.脊索动物的三大主要特征是什么?试各加以简略说明。
2.脊索动物还有哪些次要特征?为什么说它们是次要的?
3.脊索动物门可分为几个亚门?几个纲?试扼要记述一下各亚门和各纲的特点。
4.试描述海鞘的呼吸活动和摄食过程。
5.何谓逆行变态?试以海鞘为例来加以说明。
6.尾索动物的主要特点是什么?
7.头索动物何以得名?为什么说它们是原索动物中最高等的类群?
8.简述文昌鱼的外形和体内结构。
9.文昌鱼和海鞘在消化系统上有何异同?
10.试绘制文昌鱼的血液循环路线图。
11.理解和掌握文昌鱼的胚胎发育各阶段的特征。
本文标题:第十四章 脊索动物门(Chordata)
手机页面:http://m.dljs.net/dlsk/dongwuxue/59419.html
本文地址:http://www.dljs.net/dlsk/dongwuxue/59419.html
