ЕкЫФНк жВЮяЕФзщжЏКЭзщжЏЯЕЭГ
ЁЁ
ЁЁЁЁЯИАћЗжЛЏЕМжТжВЮяЬхжааЮГЩЖржжРраЭЕФЯИАћЃЌетвВОЭЪЧЯИАћЗжЛЏЕМжТСЫзщжЏЕФаЮГЩЁЃШЫУЧвЛАуАбдкИіЬхЗЂг§жаЃЌОпгаЯрЭЌРДдДЕФЃЈМДгЩЭЌвЛИіЛђЭЌвЛШКЗжЩњЯИАћЩњГЄЁЂЗжЛЏЖјРДЕФЃЉЭЌвЛРраЭЃЌЛђВЛЭЌРраЭЕФЯИАћШКзщГЩЕФНсЙЙКЭЙІФмЕЅЮЛЃЌГЦЮЊзщжЏЃЈtissueЃЉЁЃгЩвЛжжРраЭЯИАћЙЙГЩЕФзщжЏЃЌГЦМђЕЅзщжЏЃЈsimple tissueЃЉЁЃгЩЖржжРраЭЯИАћЙЙГЩЕФзщжЏЃЌГЦИДКЯзщжЏЃЈcompoundtissueЃЉЁЃ
ЁЁЁЁжВЮяУПвЛРрЦїЙйЖМАќКЌгавЛЖЈжжРрЕФзщжЏЃЌЦфжаУПвЛжжзщжЏОпгавЛЖЈЕФЗжВМЙцТЩКЭааЪЙвЛжжжївЊЕФЩњРэЙІФмЃЌЕЋЪЧетаЉзщжЏЕФЙІФмгжЪЧБиаыЯрЛЅвРРЕКЭЯрЛЅХфКЯЕФЃЌР§ШчвЖЪЧжВЮяНјааЙтКЯзїгУЕФЦїЙйЃЌЦфжажївЊЗжЛЏЮЊДѓСПЕФЭЌЛЏзщжЏНјааЙтКЯзїгУЃЌЕЋдкЫќЕФжмЮЇИВИЧзХБЃЛЄзщжЏЃЌвдЗРжЙЭЌЛЏзщжЏЖЊЪЇЫЎЗжКЭЛњаЕЫ№ЩЫЃЌДЫЭтЃЌЪфЕМзщжЏЙсДЉгкЭЌЛЏзщжЏжаЃЌБЃжЄЫЎЗжЕФЙЉгІКЭАбЭЌЛЏВњЮядЫЪфГіШЅЃЌетбљЃЌШ§жжзщжЏЯрЛЅХфКЯЃЌБЃжЄСЫвЖЕФЙтКЯзїгУе§ГЃНјааЁЃгЩДЫПЩМћЃЌзщГЩЦїЙйЕФВЛЭЌзщжЏЃЌБэЯжЮЊећЬхЬѕМўЯТЕФЗжЙЄазїЃЌЙВЭЌБЃжЄЦїЙйЙІФмЕФЭъГЩЁЃ
ЁЁ
вЛЁЂжВЮязщжЏЕФРраЭ
ЁЁ
ЁЁЁЁжВЮязщжЏЗжГЩЗжЩњзщжЏЃЈmeristematic tissueЛђmeristemЃЉКЭГЩЪьзщжЏЃЈmature tissueЃЉСНДѓРрЁЃ
ЁЁЁЁЃЈвЛЃЉЗжЩњзщжЏ
ЁЁЁЁ1ЃЎЗжЩњзщжЏЕФИХФю жжзгжВЮяжаОпЗжСбФмСІЕФЯИАћЯожЦдкжВЮяЬхЕФФГаЉВПЮЛЃЌетаЉВПЮЛЕФЯИАћдкжВЮяЬхЕФвЛЩњжаГжајЕиБЃГжЧПСвЕФЗжСбФмСІЃЌвЛЗНУцВЛЖЯдіМгаТЯИАћЕНжВЮяЬхжаЃЌСэвЛЗНУцздМКМЬај“гРДц”ЯТШЅЃЌетжжОпГжајЗжСбФмСІЕФЯИАћШКГЦЮЊЗжЩњзщжЏЁЃ
ЁЁЁЁ2ЃЎЗжЩњзщжЏЕФРраЭ
ЁЁЁЁЃЈ1ЃЉАДдкжВЮяЬхЩЯЕФЮЛжУЗж ИљОндкжВЮяЬхЩЯЕФЮЛжУЃЌПЩвдАбЗжЩњзщжЏЧјЗжЮЊЖЅЖЫЗжЩњзщжЏЃЈapical meristemЃЉЁЂВрЩњЗжЩњзщжЏЃЈlateral meristemЃЉКЭОгМфЗжЩњзщжЏЃЈintercalarymeristemЃЉЁЃ
ЁЁЁЁЂйЖЅЖЫЗжЩњзщжЏ ЖЅЖЫЗжЩњзщжЏЮЛгкОЅгыИљжїжсЕФКЭВржІЕФЖЅЖЫЃЈЭМ1Ѓ38ЃЉЁЃЫќУЧЕФЗжСбЛюЖЏПЩвдЪЙИљКЭОЅВЛЖЯЩьГЄЃЌВЂдкОЅЩЯаЮГЩВржІКЭвЖЃЌЪЙжВЮяЬхРЉДѓгЊбјУцЛ§ЁЃОЅЕФЖЅЖЫЗжЩњзщжЏзюКѓЛЙНЋВњЩњЩњжГЦїЙйЁЃ
ЁЁЁЁЖЅЖЫЗжЩњзщжЏЯИАћЕФЬиеїЪЧЃКЯИАћаЁЖјЕШОЖЃЌОпгаБЁБкЃЌЯИАћКЫЮЛгкжабыВЂеМгаНЯДѓЕФЬхЛ§ЃЌвКХнаЁЖјЗжЩЂЃЌдЩњжЪХЈКёЃЌЯИАћФкЭЈГЃШБЩйКѓКЌЮяЁЃ
ЁЁЁЁЂкВрЩњЗжЩњзщжЏ ВрЩњЗжЩњзщжЏЮЛгкИљКЭОЅЕФВрЗНЕФжмЮЇВПЗжЃЌППНќЦїЙйЕФБпдЕЃЈЭМ1—38ЃЉЁЃЫќАќРЈаЮГЩВуЃЈcambiumЃЉКЭФОЫЈаЮГЩВуЃЈcork cambiumЛђphellogenЃЉЁЃаЮГЩВуЕФЛюЖЏФмЪЙИљКЭОЅВЛЖЯдіДжЃЌвдЪЪгІжВЮягЊбјУцЛ§ЕФРЉДѓЁЃФОЫЈаЮГЩВуЕФЛюЖЏЪЧЪЙГЄДжЕФИљЁЂОЅБэУцЛђЪмЩЫЕФЦїЙйБэУцаЮГЩаТЕФБЃЛЄзщжЏЁЃ
ЁЁЁЁВрЩњЗжЩњзщжЏВЂВЛЦеБщДцдкгкЫљгажжзгжВЮяжаЃЌЫќУЧжївЊДцдкгкТузгжВЮяКЭФОБОЫЋзгвЖжВЮяжаЁЃВнБОЫЋзгвЖжВЮяжаЕФВрЩњЗжЩњзщжЏжЛгаЮЂШѕЕФЛюЖЏЛђИљБОВЛДцдкЃЌдкЕЅзгвЖжВЮяжаВрЩњЗжЩњзщжЏвЛАуВЛДцдкЃЌвђДЫЃЌВнБОЫЋзгвЖжВЮяКЭЕЅзгвЖжВЮяЕФИљКЭОЅУЛгаУїЯдЕФдіДжЩњГЄЁЃ
ЁЁЁЁВрЩњЗжЩњзщжЏЕФЯИАћгыЖЅЖЫЗжЩњзщжЏЕФЯИАћгаУїЯдЕФЧјБ№ЃЌР§ШчаЮГЩВуЯИАћДѓВПЗжГЪГЄЫѓаЮЃЌдЩњжЪЬхИпЖШвКХнЛЏЃЌЯИАћжЪВЛХЈКёЁЃЖјЧвЫќУЧЕФЗжСбЛюЖЏЭљЭљЫцМОНкЕФБфЛЏОпгаУїЯдЕФжмЦкадЁЃ
ЁЁЁЁЂлОгМфЗжЩњзщжЏ ОгМфЗжЩњзщжЏЪЧМадкЖрЩйвбОЗжЛЏСЫЕФзщжЏЧјгђжЎМфЕФЗжЩњзщжЏЃЌЫќЪЧЖЅЖЫЗжЩњзщжЏдкФГаЉЦїЙйжаОжВПЧјгђЕФБЃСєЁЃ
ЁЁЁЁЕфаЭЕФОгМфЗжЩњзщжЏДцдкгкаэЖрЕЅзгвЖжВЮяЕФОЅКЭвЖжаЃЌР§ШчЫЎЕОЁЂаЁТѓЕШКЬЙШРрзїЮяЃЌдкОЅЕФНкМфЛљВПБЃСєОгМфЗжЩњзщжЏЃЌЫљвдЕБЖЅЖЫЗжЛЏГЩгзЫыКѓЃЌШдФмНшжњгкОгМфЗжЩњзщжЏЕФЛюЖЏЃЌНјааАЮНкКЭГщЫыЃЌЪЙОЅМБОчГЄИпЃЈЭМ1Ѓ39ЃЉЁЃДаЁЂЫтЁЂОТВЫЕФвЖзгМєШЅЩЯВПЛЙФмМЬајЩьГЄЃЌетвВЪЧвђЮЊвЖЛљВПЕФОгМфЗжЩњзщжЏЛюЖЏЕФНсЙћЁЃТфЛЈЩњгЩгкДЦШяБњЛљВПОгМфЗжЩњзщжЏЕФЛюЖЏЃЌЖјФмАбПЊЛЈКѓЕФзгЗПЭЦШыЭСжаЁЃ
ЁЁЁЁОгМфЗжЩњзщжЏгыЖЅЖЫЗжЩњзщжЏКЭВрЩњЗжЩњзщжЏЯрБШЃЌЯИАћГжајЛюЖЏЕФЪБМфНЯЖЬЃЌЗжСбвЛЖЮЪБМфКѓЃЌЫљгаЕФЯИАћЖМЭъШЋзЊБфГЩГЩЪьзщжЏЁЃ

 ЁЁ
ЁЁ
ЁЁЁЁЃЈ2ЃЉАДРДдДЕФаджЪЗж ЗжЩњзщжЏвВПЩИљОнзщжЏРДдДЕФаджЪЛЎЗжЮЊдЗжЩњзщжЏ(promeri-stemЃЉЁЂГѕЩњЗжЩњзщжЏЃЈprimary meristemЃЉКЭДЮЩњЗжЩњзщжЏЃЈsecondary meristemЃЉЁЃ
ЁЁЁЁЂйдЗжЩњзщжЏ дЗжЩњзщжЏЪЧжБНггЩХпЯИАћБЃСєЯТРДЕФЃЌвЛАуОпгаГжОУЖјЧПСвЕФЗжСбФмСІЃЌЮЛгкИљЖЫКЭОЅЖЫНЯЧАЕФВПЗжЁЃ
ЁЁЁЁЂкГѕЩњЗжЩњзщжЏ ГѕЩњЗжЩњзщжЏЪЧгЩдЗжЩњзщжЏИебмЩњЕФЯИАћзщГЩЃЌетаЉЯИАћдкаЮЬЌЩЯвбГіЯжСЫзюГѕЕФЗжЛЏЃЌЕЋЯИАћШдОпгаКмЧПЕФЗжСбФмСІЃЌвђДЫЃЌЫќЪЧвЛжжБпЗжСбЁЂБпЗжЛЏЕФзщжЏЃЌвВПЩПДзїЪЧгЩЗжЩњзщжЏЯђГЩЪьзщжЏЙ§ЖЩЕФзщжЏЁЃ
ЁЁЁЁЂлДЮЩњЗжЩњзщжЏ ДЮЩњЗжЩњзщжЏЪЧгЩГЩЪьзщжЏЕФЯИАћЃЌОРњЩњРэКЭаЮЬЌЩЯЕФБфЛЏЃЌЭбРыдРДЕФГЩЪьзДЬЌЃЈМДЗДЗжЛЏЃЉЃЌжиаТзЊБфЖјГЩЕФЗжЩњзщжЏЁЃ
ЁЁЁЁШчЙћАбЖўжжЗжРрЗНЗЈЖдгІЦ№РДПДЃЌдђЙувхЕФЖЅЖЫЗжЩњзщжЏАќРЈдЗжЩњзщжЏКЭГѕЩњЗжЩњзщжЏЃЌЖјВрЩњЗжЩњзщжЏвЛАуНВЪЧЪєгкДЮЩњЗжЩњзщжЏРраЭЃЌЦфжаФОЫЈаЮГЩВуЪЧЕфаЭЕФДЮЩњЗжЩњзщжЏЁЃ
ЁЁЁЁЃЈЖўЃЉГЩЪьзщжЏ
ЁЁЁЁ1ЃЎГЩЪьзщжЏЕФИХФю ЗжЩњзщжЏбмЩњЕФДѓВПЗжЯИАћЃЌж№НЅЩЅЪЇЗжСбЕФФмСІЃЌНјвЛВНЩњГЄКЭЗжЛЏЃЌаЮГЩЕФЦфЫћИїжжзщжЏЃЌГЦЮЊГЩЪьзщжЏЃЌгаЪБвВГЦЮЊгРОУзщжЏЃЈpermanent tissueЃЉЁЃ
ЁЁЁЁИїжжГЩЪьзщжЏПЩвдОпгаВЛЭЌЕФЗжЛЏГЬЖШЃЌгааЉзщжЏЕФЯИАћгыЗжЩњзщжЏЕФВювьМЋаЁЃЌОпгавЛАуЕФДњаЛЛюЖЏЃЌВЂЧввВФмНјааЗжСбЁЃЖјСэвЛаЉзщжЏЕФЯИАћдђгаКмДѓЕФаЮЬЌИФБфЃЌЙІФмзЈвЛЃЌВЂЧвЭъШЋЩЅЪЇЗжСбФмСІЁЃвђДЫЃЌзщжЏЕФ“ГЩЪь”Лђ“гРОУ”ГЬЖШЪЧЯрЖдЕФЁЃЖјЧвГЩЪьзщжЏвВВЛЪЧвЛГЩВЛБфЕФЃЌгШЦфЪЧЗжЛЏГЬЖШНЯЧГЕФзщжЏЃЌгаЪБФмЫцзХжВЮяЕФЗЂг§ЃЌНјвЛВНЬиЛЏЮЊСэвЛРрзщжЏЃЛЯрЗДЃЌгаЪБдквЛЖЈЕФЬѕМўЯТЃЌгжПЩвдЗДЗжЛЏЃЈЛђЭбЗжЛЏdedifferentiationЃЉГЩЗжЩњзщжЏЁЃ
ЁЁЁЁ2ЃЎГЩЪьзщжЏЕФРраЭ ГЩЪьзщжЏПЩвдАДееЙІФмЗжЮЊБЃЛЄзщжЏЃЈprotective tissueЃЉЁЂБЁБкзщжЏЃЈparenchymaЃЉЁЂЛњаЕзщжЏЃЈmechanical tissueЃЉЁЂЪфЕМзщжЏЃЈconducting tissueЃЉКЭЗжУкНсЙЙЃЈsecretory structureЃЉЁЃ
ЁЁЁЁЃЈ1ЃЉБЃЛЄзщжЏ БЃЛЄзщжЏЪЧИВИЧгкжВЮяЬхБэЦ№БЃЛЄзїгУЕФзщжЏЃЌЫќЕФзїгУЪЧМѕЩйЬхФкЫЎЗжЕФеєЬкЃЌПижЦжВЮягыЛЗОГЕФЦјЬхНЛЛЛЃЌЗРжЙВЁГцКІЧжЯЎКЭЛњаЕЫ№ЩЫЕШЁЃБЃЛЄзщжЏАќРЈБэЦЄЃЈepidermisЃЉКЭжмЦЄЃЈperidermЃЉЁЃ
ЁЁЁЁЂйБэЦЄ БэЦЄгжГЦБэЦЄВуЃЌЪЧгзФлЕФИљКЭОЅЁЂвЖЁЂЛЈЁЂЙћЪЕЕШЕФБэУцВуЯИАћЁЃЫќЪЧжВЮяЬхгыЭтНчЛЗОГЕФжБНгНгДЅВуЃЌвђДЫЃЌЫќЕФЬиЕугыетвЛЬиЪтЮЛжУКЭЩњРэЙІФмУмЧагаЙиЁЃ
ЁЁЁЁБэЦЄвЛАужЛгавЛВуЯИАћЃЌЕЋЫќВЛжЛЪЧгЩвЛРрЯИАћзщГЩЃЌЭЈГЃКЌгаЖржжВЛЭЌЬиеїКЭЙІФмЕФЯИАћЃЌЦфжаБэЦЄЯИАћЪЧзюЛљБОЕФГЩЗжЃЌЦфЫћЯИАћЗжЩЂгкБэЦЄЯИАћжЎМфЁЃ
ЁЁЁЁБэЦЄЯИАћГЪИїжжаЮзДЕФАхПщзДЃЌХХСаЪЎЗжНєУмЃЌГ§ЦјПзЭтЃЌВЛДцдкСэЭтЕФЯИАћМфЯЖЁЃБэЦЄЯИАћЪЧЩњЛюЯИАћЃЌЯИАћвЛАуВЛОпвЖТЬЬхЃЌЕЋГЃгаАзЩЋЬхКЭгаЩЋЬхЃЌЯИАћФкДЂВигаЕэЗлСЃКЭЦфЫћДњаЛВњЮяШчЩЋЫиЁЂЕЄФўЁЂОЇЬхЕШЁЃОЅКЭвЖЕШжВЮяЬхЦјЩњВПЗжЕФБэЦЄЯИАћЃЌЭтЯвЯђБкЭљЭљНЯКёЃЌВЂНЧжЪЛЏЃЌДЫЭтЃЌдкБкЕФБэУцЛЙГСЛ§вЛВуУїЯдЕФНЧжЪВуЃЌЪЙБэЦЄОпгаИпЖШЕФВЛЭИЫЎадЃЌгааЇЕиМѕЩйСЫЬхФкЕФЫЎЗжеєЬкЃЌМсгВЕФНЧжЪВуЖдЗРжЙВЁОњЕФЧжШыКЭдіМгЛњаЕжЇГжЃЌвВгавЛЖЈЕФзїгУЁЃгааЉжВЮяЃЈШчИЪесЕФОЅЁЂЦЯЬбЁЂЦЛЙћЕФЙћЪЕЃЉдкНЧжЪВуЭтЛЙОпгавЛВуРЏжЪЕФ“ЫЊ”ЃЌЫќЕФзїгУЪЧЪЙБэУцВЛвзНўЪЊЃЌОпгаЗРжЙВЁОњцпзгдкЬхБэУШЗЂЕФзїгУЁЃдкЩњВњЪЕМљжаЃЌжВЮяЬхБэУцВуЕФНсЙЙЧщПіЃЌЪЧбЁг§ПЙВЁЦЗжжЃЌЪЙгУХЉвЉЛђГ§ВнМСЪББиаыПМТЧЕФвђЫиЁЃБэЦЄЕФНсЙЙКЭНЧжЪВуЮЦаЭЃЌвВЪЧжВЮяЗжРрЩЯЕФвЛИівРОнЃЈЭМ1Ѓ40ЃЌAЁЂBЃЛЭМ1Ѓ41ЃЉ

 ЁЁ
ЁЁ
ЁЁЁЁЯждкЭЈЙ§ЕчзгЯдЮЂОЕЕФЙлВьбаОПЃЌЖдНЧжЪВуЕФНсЙЙгаСЫНјвЛВНЕФСЫНтЃЌЫќАќРЈЖўВуЃЌЮЛгкЭтУцЕФвЛВугЩНЧжЪКЭРЏжЪзщГЩЃЌЮЛгкРяУцЕФвЛВугЩНЧжЪКЭЯЫЮЌЫизщГЩЁЃгаШЫЬсГіНЋетЖўВуКЯГЦЮЊНЧжЪФЄЃЈЯрЕБгкдРДЕФНЧжЪВуЃЉЃЌЖјНЋЭтВуГЦЮЊНЧжЪВуЃЌНЋФкВуГЦЮЊНЧЛЏВуЁЃНЧЛЏВуКЭГѕЩњБкжЎМфУїЯдЕигаЙћНКВуЗжНчЃЈЭМ1Ѓ40ЃЌCЃЉЁЃ
ЁЁЁЁдкЦјЩњБэЦЄЩЯОпгааэЖрЦјПзЃЈstomaЃЉЃЌЫќУЧЪЧЦјЬхГіШыжВЮяЬхЕФУХЛЇЁЃЦјПзЪЧгЩЖўИіЬиЪтЕФЯИАћМДБЃЮРЯИАћЃЈguard cellЃЉКЭЫќУЧМфЕФПЊПкЙВЭЌзщГЩЕФЁЃдкгааЉГЁКЯЃЌвВгаЕЅАбПЊПкГЦЦјПзЕФЁЃБЃЮРЯИАћГЩЩіаЮЛђбЦСхаЮЃЌЯИАћФкКЌгавЖТЬЬхЃЌЬиЪтЕФВЛОљдШдіКёЕФЯИАћБкЃЌЪЙБЃЮРЯИАћаЮзДИФБфЪБФмЕМжТПзПкЕФПЊЗХЛђЙиБеЃЌДгЖјЕїНкЦјЬхЕФГіШыКЭЫЎЗжЕФеєЬкЃЈЭМ1Ѓ41ЃЌBЃЉЁЃ
ЁЁЁЁБэЦЄЛЙПЩвдОпгаИїжжЕЅЯИАћЛђЖрЯИАћЕФУЋзДИНЪєЮяЃЈЭМ1—42ЃЉЁЃвЛАуШЯЮЊБэЦЄУЋОпгаБЃЛЄКЭЗРжЙЯТУцЫЎЗжЩЅЪЇЕФзїгУЁЃЮвУЧгУЕФУоКЭФОУоЯЫЮЌЃЌЖМЪЧЫќУЧжжЦЄЩЯЕФБэЦЄУЋЁЃгааЉжВЮяОпгаЗжУкЙІФмЕФБэЦЄУЋЃЌПЩвдЗжУкГіЗМЯугЭЁЂеГвКЁЂЪїжЌЁЂеСФдЕШЮяжЪЁЃ
ЁЁЁЁИљЕФБэЦЄжївЊгыЮќЪеЫЎЗжКЭЮоЛњбЮгаЙиЃЌвђДЫЃЌЫќЪЧвЛжжЮќЪезщжЏЃЈabsorptive tissueЃЉЁЃИљЕФБэЦЄЯИАћОпгаБЁЕФБкКЭБЁЕФНЧжЪВуЃЌаэЖрЯИАћЕФЭтБкЯђЭтбгЩьЃЌаЮГЩЯИГЄЕФЙмзДЭЛЦ№——ИљУЋЃЈroot hairЃЉЃЌДгЖјДѓДѓЕигаРћгкИљЕФЮќЪеЁЃ
ЁЁЁЁБэЦЄдкжВЮяЬхЩЯДцдкЕФЪБМфЃЌвРЫљдкЦїЙйЪЧЗёОпгаМгДжЩњГЄЖјвьЃЌОпгаУїЯдМгДжЩњГЄЕФЦїЙйЃЌШчТузгжВЮяКЭДѓВПЗжЫЋзгвЖжВЮяЕФИљКЭОЅЃЌБэЦЄЛсвђЦїЙйЕФдіДжЖјЦЦЛЕЁЂЭбТфЃЌгЩФкЗНВњЩњЕФДЮЩњБЃЛЄзщжЏ——жмЦЄЫљШЁДњЁЃдкНЯЩйЛђУЛгаДЮЩњЩњГЄЕФЦїЙйЩЯЃЌР§ШчвЖЁЂЙћЪЕЁЂДѓВПЗжЕЅзгвЖжВЮяЕФИљКЭОЅЩЯЃЌБэЦЄПЩГЄЦкДцдкЁЃ
ЁЁЁЁЂкжмЦЄ жмЦЄЪЧШЁДњБэЦЄЕФДЮЩњБЃЛЄзщжЏЃЌДцдкгкгаМгДжЩњГЄЕФИљКЭОЅЕФБэУцЁЃЫќгЩВрЩњЗжЩњзщжЏ——ФОЫЈаЮГЩВуаЮГЩЁЃФОЫЈаЮГЩВуЦНжмЕиЗжСбЃЌаЮГЩОЖЯђГЩааЕФЯИАћааСаЃЌетаЉЯИАћЯђЭтЗжЛЏГЩФОЫЈЃЈphellemЛђcorkЃЉЃЌЯђФкЗжЛЏГЩЫЈФкВуЃЈphellodermЁЃФОЫЈЁЂФОЫЈаЮГЩВуКЭЫЈФкВуКЯГЦжмЦЄЃЈЭМ1Ѓ43ЃЉЁЃ

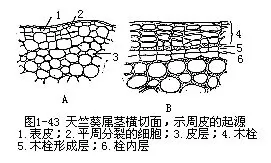 ЁЁ
ЁЁ
ЁЁЁЁФОЫЈОпЖрВуЯИАћЃЌдкКсЧаУцжаЯИАћГЪГЄЗНаЮЃЌНєУмХХСаГЩећЦыЕФОЖЯђааСаЃЌЯИАћБкНЯКёЃЌВЂЧвЧПСвЫЈЛЏЃЌЯИАћГЩЪьЪБдЩњжЪЬхЫРЭіНтЬхЃЌЯИАћЧЛФкЭЈГЃГфТњПеЦјЁЃетаЉЬиеїЪЙФОЫЈОпгаИпЖШВЛЭИЫЎадЃЌВЂгаПЙбЙЁЂИєШШЁЂОјдЕЁЂжЪЕиЧсЁЂОпЕЏадЁЂПЙгаЛњШмМСКЭЖржжЛЏбЇвЉЦЗЕФЬиадЃЌЖджВЮяЬхЦ№СЫгааЇЕФБЃЛЄзїгУЁЃЭЌЪБвВЪЙЫќдкЩЬвЕЩЯгаЯрЕБЕФживЊадЃЌПЩЙЉШегУЛђзїЧсжЪОјдЕВФСЯКЭОШЩњЩшБИЕШЁЃЫЈЦЄДЂЁЂЫЈЦЄшнКЭЛЦщоЃЈЛЦАиЃЉЪЧЩЬгУФОЫЈЕФжївЊРДдДЁЃ
ЁЁЁЁЫЈФкВуЪЧБЁБкЕФЩњЛюЯИАћЃЌГЃГЃжЛгавЛВуЯИАћКёЃЌвЛАужЛФмДгЫќУЧгыЭтУцЕФФОЫЈЯИАћХХГЩЭЌвЛећЦыЕФОЖЯђааСаЃЌЖјгыЦЄВуБЁБкЯИАћЧјБ№ПЊРДЁЃ
ЁЁЁЁдкжмЦЄЕФФГаЉЯоЖЈВПЮЛЃЌЦфФОЫЈаЮГЩВуЯИАћБШЦфЫћВПЗжИќЮЊЛюдОЃЌЯђЭтбмЩњГівЛжжгыФОЫЈЯИАћВЛЭЌЃЌВЂОпгаЗЂДяЯИАћМфЯЖЕФзщжЏЃЈВЙГфзщжЏЃЉЁЃЫќУЧЭЛЦЦжмЦЄЃЌдкЪїЦЄБэУцаЮГЩИїжжаЮзДЕФаЁЭЛЦ№ЃЌГЦЮЊЦЄПзЃЈlenticelЃЉЁЃЦЄПзЪЧжмЦЄЩЯЕФЭЈЦјНсЙЙЃЌЮЛгкжмЦЄФкЕФЩњЛюЯИАћЃЌФмЭЈЙ§ЫќУЧгыЭтНчНјааЦјЬхНЛЛЛЁЃ
ЁЁЁЁЃЈ2ЃЉБЁБкзщжЏ БЁБкзщжЏЪЧНјааИїжжДњаЛЛюЖЏЕФжївЊзщжЏЃЌЙтКЯзїгУЁЂКєЮќзїгУЁЂжќВизїгУМАИїРрДњаЛЮяЕФКЯГЩКЭзЊЛЏЖМжївЊгЩЫќНјааЁЃБЁБкзщжЏеМжВЮяЬхЬхЛ§ЕФДѓВПЗжЃЌШчОЅКЭИљЕФЦЄВуМАЫшВПЁЂвЖШтЯИАћЁЂЛЈЕФИїВПЃЌаэЖрЙћЪЕКЭжжзгжаЃЌШЋВПЛђжївЊЪЧБЁБкзщжЏЃЌЦфЫћЖржжзщжЏЃЌШчЛњаЕзщжЏКЭЪфЕМзщжЏЕШЃЌГЃАќТёгкЦфжаЁЃвђДЫЃЌДгФГжжвтвхНВЃЌБЁБкзщжЏЪЧжВЮяЬхзщГЩЕФЛљДЁЃЌвВОЭЪЧЛљБОзщжЏЕФжївЊзщГЩВПЗжЃЌДЫЭтЃЌЛљБОзщжЏЭЈГЃЛЙАќРЈКёНЧзщжЏКЭКёБкзщжЏЁЃ
ЁЁЁЁБЁБкзщжЏвдЯИАћОпгаБЁЕФГѕЩњБкЖјЕУУћЃЌЫќЪЧвЛРрНЯВЛЗжЛЏЕФГЩЪьзщжЏЁЃДгНсЙЙЩЯПДЃЌБЁБкзщжЏНЯЩйЬиЛЏЖјНЯЖрЕиНгНќЗжЩњзщжЏЃЌГ§СЫЯИАћБкЖўепЖМЪЧБЁЕФГѕЩњБкЭтЃЌЯИАћЕФаЮзДврГЃГЪЕШОЖЕФЖрУцЬхЃЌЯИАћОпгаЩњЛюЕФдЩњжЪЬхЁЃДгЗжЩњФмСІЩЯПДЃЌБЁБкзщжЏврФмНјаагаЯоЕФЗжСбЁЃдкДДЩЫгњКЯЁЂдйЩњзїгУаЮГЩВЛЖЈИљКЭВЛЖЈбПЃЌвдМАМоНггњКЯЕШЪБЦкБЁБкзщжЏЯИАћФмЗЂЩњЗДЗжЛЏЃЌзЊБфЮЊЗжЩњзщжЏЁЃдке§ГЃзДЬЌЯТЃЌЫќУЧвВВЮгыВрЩњЗжЩњзщжЏЕФЗЂЩњЁЃгЩДЫПЩМћЃЌБЁБкзщжЏгазХКмЧПЕФЗжЩњЧБФмЃЌдквЛЖЈЬѕМўЯТЃЌКмШнвззЊЛЏЮЊЗжЩњзщжЏЁЃДЫЭтЃЌБЁБкзщжЏгаНЯДѓЕФПЩЫмадЃЌдкжВЮяЬхЗЂг§ЕФЙ§ГЬжаЃЌГЃФмНјвЛВНЗЂг§ЮЊЬиЛЏГЬЖШИќИпЕФзщжЏЃЌШчЗЂг§ЮЊКёБкзщжЏЁЃБЁБкзщжЏЕФСэвЛЬиЕуЃЌЪЧвЛАуЖМОпгаНЯЗЂДяЕФЯИАћМфЯЖЃЌетЖдгкЯИАћЕФЭњЪЂДњаЛЪЧБиашЕФЁЃ
ЁЁЁЁБЁБкзщжЏвђЙІФмВЛЭЌПЩЗжГЩВЛЭЌЕФРраЭЃЌЫќУЧдкаЮЬЌЩЯгаИїздЕФЬиЕуЃЈЭМ1-44ЃЉЁЃ

ЁЁ
ЁЁЁЁгЊЙтКЯзїгУЕФБЁБкзщжЏГЦЮЊЭЌЛЏзщжЏЃЈassimilating tissueЃЉЃЌжївЊЬиЕуЪЧдЩњжЪЬхжаЗЂг§ГіДѓСПЕФвЖТЬЬхЁЃЭЌЛЏзщжЏЗжВМгкжВЮяЬхЕФвЛЧаТЬЩЋВПЗжЃЌШчгзОЅЕФЦЄВуЃЌЗЂг§жаЕФЙћЪЕКЭжжзгжаЃЌгШЦфЪЧвЖЕФвЖШтЃЌЪЧЕфаЭЕФЭЌЛЏзщжЏЁЃ
ЁЁЁЁжќВиДѓСПгЊбјЮяжЪЕФБЁБкзщжЏЃЌГЦЮЊДЂВизщжЏЃЈstorage tissueЃЉЁЃжївЊДцдкгкИїРржќВиЦїЙйЃЌШчПщИљЁЂПщОЅЁЂЧђОЅЁЂСлОЅЁЂЙћЪЕКЭжжзгжаЃЌИљЁЂОЅЕФЦЄВуКЭЫшЃЌвдМАЦфЫћБЁБкзщжЏвВЖМОпгажќВиЕФЙІФмЁЃ
ЁЁЁЁжќВигаЗсИЛЫЎЗжЕФЯИАћЃЌГЦЮЊДЂЫЎзщжЏЃЈaqueous tissueЃЉЁЃЫќЕФЯИАћНЯДѓЃЌвКХнжаКЌгаДѓСПЕФеГаджвКЁЃвЛАуДцдкгкКЕЩњЕФШтжЪжВЮяжаЃЌШчЯЩШЫеЦЁЂСњЩрРМЁЂОАЬьЁЂТЋміЕШЕФЙтКЯЦїЙйжаЖМФмПДЕНЁЃ

ЁЁ
ЁЁЁЁОпгаДѓСПЯИАћМфЯЖЕФБЁБкзщжЏЃЌГЦЮЊЭЈЦјзщжЏЃЈaerenchymaЃЉЁЃдкЫЎЩњКЭЪЊЩњжВЮяжаЃЌДЫРрзщжЏЬиБ№ЗЂДяЃЌШчЫЎЕОЁЂСЋЁЂЫЏСЋЕШЕФИљЁЂОЅЁЂвЖжаБЁБкзщжЏгаДѓЕФМфЯЖЃЌдкЬхФкаЮГЩвЛИіЯрЛЅЙсЭЈЕФЭЈЦјЯЕЭГЃЌЪЙвЖгЊЙтКЯзїгУЖјВњЩњЕФбѕЦјФмЭЈЙ§ЫќНјШыИљжаЁЃЭЈЦјзщжЏЛЙгыдкЫЎжаЕФИЁСІКЭжЇГжзїгУгаЙиЁЃ
ЁЁЁЁ60ФъДњЃЌдЫгУЕчзгЯдЮЂОЕаТЗЂЯжвЛРрЬиЛЏЕФБЁБкЯИАћЁЃетжжЯИАћзюЯджјЕФЬиеїЪЧЯИАћБкОпФкЭЛЩњГЄЃЌМДЯђФкЭЛШыЯИАћЧЛФкЃЌаЮГЩаэЖржИзДЛђТЙНЧзДЕФВЛЙцдђЭЛЦ№ЁЃетбљЪЙЕУНєЬљдкБкФкВрЕФжЪФЄУцЛ§ДѓДѓдіМгЃЌРЉДѓСЫдЩњжЪЬхЕФБэУцЛ§гыЬхЛ§жЎБШЃЌДгЖјгаРћгкЯИАћДгжмЮЇбИЫйЕиЮќЪеЮяжЪЃЌвВгаРћгкЮяжЪбИЫйЕиДгдЩњжЪЬхжаЪЭЗХГіШЅЁЃЫљвдЃЌетаЉЯИАћЪЧвЛРргыЮяжЪбИЫйЕиДЋЕнУмЧаЯрЙиЕФБЁБкЯИАћЃЌЬиГЦЮЊДЋЕнЯИАћЃЈtransfer cellЃЉЃЌвВГЦзЊЪфЯИАћЛђзЊвЦЯИАћЃЈЭМ1Ѓ45ЃЉЁЃЫќУЧдкжВЮяЬхФкЃЌЖМЪЧГіЯждкШмжЪЖЬЭОУмМЏдЫЪфЕФВПЮЛЃЌР§ШчЦеБщДцдкгквЖЕФаЁвЖТіжаЃЌдкЪфЕМЗжзгжмЮЇЃЌГЩЮЊвЖШтКЭЪфЕМЗжзгжЎМфЮяжЪдЫЪфЕФЧХСКЁЃдкаэЖржВЮяОЅЛђЛЈађжсНкВПЕФЮЌЙмзщжЏжаЃЌдкЗжУкНсЙЙжаЃЌдкжжзгЕФзгвЖЁЂХпШщЛђХпБњЕШВПЮЛвВгаЗжВМЁЃДЋЕнЯИАћЪЧЛюЯИАћЃЌЯИАћБквЛАуЮЊГѕЩњБкЃЌАћМфСЌЫПЗЂДяЃЌЯИАћКЫаЮзДЖрбљЃЌЦфЫћШчЯпСЃЬхЁЂИпЖћЛљЬхЁЂКЫЬЧЬхЁЂЮЂЬхЕШвВЖМБШНЯЗсИЛЁЃДЋЕнЯИАћЕФЗЂЯжЪЙШЫУЧЖдЮяжЪдкЩњЛюЯИАћМфЕФИпаЇТЪЕФдЫЪфКЭДЋЕнгаСЫИќНјвЛВНЕФШЯЪЖЁЃ
ЁЁЁЁЃЈ3ЃЉЛњаЕзщжЏ ЛњаЕзщжЏЪЧЖджВЮяЦ№жївЊжЇГжзїгУЕФзщжЏЁЃЫќгаКмЧПЕФПЙбЙЁЂПЙеХКЭПЙЧњФгЕФФмСІЃЌжВЮяФмгавЛЖЈЕФгВЖШЃЌжІИЩФмЭІСЂЃЌЪївЖФмЦНеЙЃЌФмОЪмПёЗчБЉгъМАЦфЫћЭтСІЕФЧжЯЎЃЌЖМгыетжжзщжЏЕФДцдкгаЙиЁЃ
ЁЁЁЁИљОнЯИАћНсЙЙЕФВЛЭЌЃЌЛњаЕзщжЏПЩЗжЮЊКёНЧзщжЏЃЈcollenchymaЃЉКЭКёБкзщжЏЃЈsclerencny-maЃЉЖўРрЁЃ
ЁЁЁЁЂйКёНЧзщжЏ КёНЧзщжЏЯИАћзюУїЯдЕФЬиеїЪЧЯИАћБкОпгаВЛОљдШЕФдіКёЃЌЖјЧветжждіКёЪЧГѕЩњБкаджЪЕФЁЃБкЕФдіКёЭЈГЃдкМИИіЯИАћСкНгДІЕФНЧИєЩЯЬиБ№УїЯдЃЌЙЪГЦКёНЧзщжЏЃЈЭМ1—46ЃЉЁЃЕЋвВгааЉжВЮяЕФКёНЧзщжЏЪЧЯИАћЕФЯвЯђБкЬиБ№КёЁЃ

ЁЁ
ЁЁЁЁКёНЧЯИАћБкЕФЛЏбЇГЩЗжЃЌГ§ЯЫЮЌЫиЭтЃЌЛЙКЌгаДѓСПЕФЙћНККЭАыЯЫЮЌЫиЃЌВЛКЌФОжЪЁЃгЩгкЙћНКгаЧПСвЕФЧзЫЎадЃЌвђДЫЃЌБкжаКЌгаДѓСПЕФЫЎЗжЃЌдкЙтбЇЯдЮЂОЕЯТЃЌдіКёЕФБкЯдГіЬиЪтЕФжщЙтЃЌКмШнвзгыЦфЫћзщжЏЯрЧјБ№ЁЃЕЋЕБжЦГЩгРОУЧаЦЌЪБЃЌВФСЯвЛОЭбЫЎЃЌдіКёЕФБкЛсБфБЁЃЌЭЌЪБжщЙтвВЛсЯћЪЇЁЃ
ЁЁЁЁКёНЧзщжЏгыБЁБкзщжЏОпгааэЖрЯрЫЦадЃЌГ§ЯИАћБкЕФГѕЩњаджЪЭтЃЌКёНЧзщжЏвВЪЧЩњЛюЯИАћЃЌвВОГЃЗЂг§ГівЖТЬЬхЃЌЯИАћврОпгаЗжСбЕФЧБФмЃЌдкаэЖржВЮяжаЃЌЫќУЧФмВЮгыФОЫЈаЮГЩВуЕФаЮГЩЁЃвђДЫЃЌвВгаШЫНЋЫќЙщРргкЬиЪтЕФБЁБкзщжЏЁЃ

ЁЁ
ЁЁЁЁКёНЧзщжЏЗжВМгкОЅЁЂвЖБњЁЂвЖЦЌЁЂЛЈБњЕШВПЗжЃЌИљжавЛАуВЛДцдкЁЃКёНЧзщжЏЕФЗжВМОпгавЛИіУїЯдЕФЬиеїЃЌМДвЛАузмЪЧЗжВМгкЦїЙйЕФЭтЮЇЃЌЛђжБНгдкБэЦЄЯТЃЌЛђгыБэЦЄжЛИєПЊМИВуБЁБкЯИАћЁЃдкОЅКЭвЖБњжаКёНЧзщжЏЭљЭљГЩСЌајЕФдВЭВЛђЗжРыГЩЪјЃЌГЃдкОпМЙзДЭЛЦ№ЕФОЅКЭвЖБњжаРтЕФВПЗжЬиБ№ЗЂДяЃЌР§ШчдкБЁКЩЕФЗНОЅжаЃЌФЯЙЯЁЂЧлВЫОпРтЕФОЅКЭвЖБњжаЁЃдквЖЦЌжаЃЌКёНЧзщжЏГЩЪјЕиЮЛгкНЯДѓвЖТіЕФвЛВрЛђЖўВрЃЈЭМ1—47ЃЉЁЃ
ЁЁЁЁКёНЧзщжЏжївЊЪЧе§дкЩњГЄЕФОЅКЭвЖЕФжЇГжзщжЏЃЌвЛЗНУцгЩгкКёНЧЯИАћЮЊГЄжљаЮЃЌЯрЛЅжиЕўХХСаЃЌГѕЩњБкЫфШЛБШНЯШэЃЌЕЋаэЖрЯИАћБкЕФдіКёВПЗжМЏжадквЛЦ№аЮГЩжљзДЛђАхзДЃЌвђЖјЪЙЫќгаНЯЧПЕФЛњаЕЧПЖШЃЛСэвЛЗНУцКёНЧзщжЏЗжЛЏНЯдчЃЌЕЋБкЕФГѕЩњаджЪЪЙЫќФмЫцзХжмЮЇЯИАћЕФбгЩьЖјРЉеЙЁЃвђДЫЃЌЫќМШгажЇГжзїгУЃЌгжВЛЗСАгзФлЦїЙйЕФЩњГЄЁЃДѓВПЗжжВЮяЕФОЅКЭвЖБњдкМЬајЗЂг§ЪБЃЌдкНЯЩюШыЕФВПЮЛгжЗЂг§ГіКёБкзщжЏЃЌетЪБЃЌКёНЧзщжЏЕФжЇГжзїгУБуГЩЮЊДЮвЊЕФСЫЁЃдкаэЖрВнжЪОЅКЭвЖжаЃЌШчВЛВњЩњКмЖрКёБкзщжЏЪБЃЌКёНЧзщжЏОЭФмМЬајГЩЮЊжївЊЕФжЇГжзщжЏЁЃгаЪБКёНЧзщжЏФмНјвЛВНЗЂг§ГіДЮЩњБкВЂФОжЪЛЏЃЌзЊБфГЩКёБкзщжЏЁЃ
ЁЁЁЁЂкКёБкзщжЏ КёБкзщжЏгыКёНЧзщжЏВЛЭЌЃЌЯИАћОпгаОљдШдіКёЕФДЮЩњБкЃЌВЂЧвГЃГЃФОжЪЛЏЁЃЯИАћГЩЪьЪБЃЌдЩњжЪЬхЭЈГЃЫРЭіЗжНтЃЌГЩЮЊжЛСєгаЯИАћБкЕФЫРЯИАћЁЃ
ЁЁЁЁИљОнЯИАћЕФаЮЬЌЃЌКёБкзщжЏПЩЗжЮЊЪЏЯИАћЃЈsclereidЛђ stone cellЃЉКЭЯЫЮЌЃЈfiberЃЉЖўРрЁЃ
ЁЁЁЁЪЏЯИАћ ЖрЮЊЕШОЖЛђТдЮЊЩьГЄЕФЯИАћЃЌгааЉОпВЛЙцдђЕФЗжжІГЩаЧУЂзДЃЌвВгаЕФНЯЯИГЄЁЃЫќУЧЭЈГЃОпгаКмКёЕФЁЂЧПСвФОжЪЛЏЕФДЮЩњБкЃЌБкЩЯгаКмЖрдВаЮЕФЕЅЮЦПзЃЌгЩгкБкЬиБ№КёЖјаЮГЩУїЯдЕФЙмзДЮЦПзЕРЃЌгаЪБЃЌЮЦПзЕРЫцБкЕФдіКёБЫДЫЛуКЯЃЌЛсаЮГЩЬиЪтЕФЗжжІЮЦПзЕРЁЃЯИАћГЩЪьЪБдЩњжЪЬхЭЈГЃЯћЪЇЃЌжЛСєЯТПеЖјаЁЕФЯИАћЧЛЃЈЭМ1Ѓ48ЃЉЁЃ

ЁЁ
ЁЁЁЁЪЏЯИАћЙуЗКЗжВМгкжВЮяЕФОЅЁЂвЖЁЂЙћЪЕКЭжжзгжаЃЌгадіМгЦїЙйЕФгВЖШКЭжЇГжЕФзїгУЁЃЫќУЧГЃГЃЕЅИіЩЂЩњЛђЪ§ИіМЏКЯГЩДиАќТёгкБЁБкзщжЏжаЃЌгаЪБвВПЩСЌајГЩЦЌЕиЗжВМЁЃР§ШчРцЙћШтжаМсгВЕФПХСЃЃЌБуЪЧГЩДиЕФЪЏЯИАћЃЌЫќУЧЪ§СПЕФЖрЩйЪЧРцЦЗжЪгХСгЕФвЛИіживЊжИБъЁЃВшЁЂЙ№ЛЈЕФвЖЦЌжаЃЌОпгаЕЅИіЕФЗжММзДЪЏЯИАћЃЌЩЂВМгквЖШтЯИАћМфЃЌдіМгСЫвЖЕФгВЖШЃЌгыВшвЖЕФЦЗжЪвВгаЙиЯЕЁЃКЫЬвЁЂЬвЁЂвЌзгЙћЪЕжаМсгВЕФКЫЃЌБуЪЧЖрВуСЌајЕФЪЏЯИАћзщГЩЕФЙћЦЄЁЃаэЖрЖЙРрЕФжжЦЄвВвђОпЖрВуЪЏЯИАћЖјБфЕУКмгВЁЃдкФГаЉжВЮяЕФОЅжавВгаГЩЖбЛђГЩЦЌЕФЪЏЯИАћЗжВМгкЦЄВуЁЂЫшЛђЮЌЙмЪјжаЁЃ
ЁЁЁЁЯЫЮЌ ЪЧЖўЖЫМтЯИГЩЫѓзДЕФЯИГЄЯИАћЃЌГЄЖШвЛАуБШПэЖШДѓаэЖрБЖЁЃЯИАћБкУїЯдЕиДЮЩњдіКёЃЌЕЋФОжЪЛЏГЬЖШКмВЛвЛжТЃЌДгВЛФОжЪЛЏЕНЧПСвФОжЪЛЏЕФЖМгаЁЃБкЩЯЮЦПзНЯЪЏЯИАћЕФЯЁЩйЃЌВЂГЃГЪЗьЯЖзДЁЃГЩЪьЪБдЩњжЪЬхвЛАуЖМЯћЪЇЃЌЯИАћЧЛГЩЮЊжаПеЃЌЩйЪ§ЯЫЮЌПЩБЃСєдЩњжЪЬхЃЌЩњЛюНЯГЄЕФвЛЖЮЪБМфЃЈЭМ1-49ЃЉЁЃ

ЁЁ
ЁЁЁЁЯЫЮЌЙуЗКЗжВМгкГЩЪьжВЮяЬхЕФИїВПЗжЁЃМтЖјЯИГЄЕФЯЫЮЌЭЈГЃдкЬхФкЯрЛЅжиЕўХХСаЃЌНєУмЕиНсКЯГЩЪјЃЌвђДЫЃЌИќдіМгзщжЏЕФЧПЖШЃЌЪЙЫќОпгаДѓЕФПЙбЙФмСІКЭЕЏадЃЌГЩЮЊГЩЪьжВЮяЬхжажївЊЕФжЇГжзщжЏЁЃ
ЁЁЁЁЃЈ4ЃЉЪфЕМзщжЏ ЪфЕМзщжЏЪЧжВЮяЬхжаЕЃИКЮяжЪГЄЭОдЫЪфЕФжївЊзщжЏЁЃИљДгЭСШРжаЮќЪеЕФЫЎЗжКЭЮоЛњбЮЃЌгЩЫќУЧдЫЫЭЕНЕиЩЯВПЗжЁЃвЖЕФЙтКЯзїгУЕФВњЮяЃЌгЩЫќУЧдЫЫЭЕНИљЁЂОЅЁЂЛЈЁЂЙћЪЕжаШЅЁЃжВЮяЬхИїВПЗжжЎМфОГЃНјааЕФЮяжЪЕФжиаТЗжХфКЭзЊвЦЃЌвВвЊЭЈЙ§ЪфЕМзщжЏРДНјааЁЃ
ЁЁЁЁдкжВЮяжаЃЌЫЎЗжЕФдЫЪфКЭгаЛњЮяЕФдЫЪфЃЌЗжБ№гЩЖўРрЪфЕМзщжЏРДГаЕЃЃЌвЛРрЮЊФОжЪВПЃЈxy-lemЃЉЃЌжївЊдЫЪфЫЎЗжКЭШмНтгкЦфжаЕФЮоЛњбЮЃЛСэвЛРрЮЊШЭЦЄВПЃЈphloemЃЉЃЌжївЊдЫЪфгаЛњгЊбјЮяжЪЁЃ
ЁЁЁЁЂйФОжЪВП ФОжЪВПЪЧгЩМИжжВЛЭЌРраЭЕФЯИАћЙЙГЩЕФвЛжжИДКЯзщжЏЃЌЫќЕФзщГЩАќКЌЙмАћЃЈtracheidЃЉКЭЕМЙмЗжзгЃЈvesselelementЛђvesselmemberЃЉЁЂЯЫЮЌЁЂБЁБкЯИАћЕШЁЃЦфжаЙмАћКЭЕМЙмЗжзгЪЧзюживЊЕФГЩдБЃЌЫЎЕФдЫЪфЪЧЭЈЙ§ЫќУЧРДЪЕЯжЕФЁЃ
ЁЁЁЁЙмАћКЭЕМЙмЗжзгЖМЪЧКёБкЕФЩьГЄЯИАћЃЌГЩЪьЪБЖМУЛгаЩњЛюЕФдЩњжЪЬхЃЌДЮЩњБкОпгаИїжжЪНбљЕФФОжЪЛЏдіКёЃЌдкБкЩЯГЪЯжГіЛЗЮЦЁЂТнЮЦЁЂЬнЮЦЁЂЭјЮЦКЭПзЮЦЕФИїжжЪНбљЃЈЭМ1—50ЃЌA—DЃЌЭМ1Ѓ51ЃЉЁЃШЛЖјЃЌЙмАћКЭЕМЙмЗжзгдкНсЙЙЩЯКЭЙІФмЩЯЪЧВЛЭъШЋЯрЭЌЕФЁЃ

 ЁЁ
ЁЁ
ЁЁЁЁЙмАћЪЧЕЅИіЯИАћЃЌФЉЖЫаЈаЮЃЌдкЦїЙйжазнЯђСЌНгЪБЃЌЩЯЁЂЯТЖўЯИАћЕФЖЫВПНєУмЕижиЕўЃЌЫЎЗжЭЈЙ§ЙмАћБкЩЯЕФЮЦПзЃЌДгвЛИіЯИАћСїЯђСэвЛИіЯИАћЁЃЙмАћДѓЖрОпНЯКёЕФБкЃЌЧвгажиЕўЕФХХСаЗНЪНЃЌЪЙЫќдкжВЮяЬхжаЛЙМцгажЇГжЕФЙІФмЁЃЫљгаЮЌЙмжВЮяЖМОпгаЙмАћЃЌЖјЧвДѓЖрЪ§оЇРржВЮяКЭТузгжВЮяЕФЪфЫЎЗжзгЃЌжЛгЩЙмАћзщГЩЁЃдкЯЕЭГЗЂг§жаЃЌЙмАћЯђЖўИіЗНЯђбнЛЏЃЌвЛИіЗНЯђЪЧЯИАћБкИќМгдіКёЃЌБкЩЯЮЦПзБфеЃЌЬиЛЏЮЊзЈгЊжЇГжЙІФмЕФФОЯЫЮЌЃЛСэвЛИіЗНЯђЪЧЯИАћЖЫБкШмНтЃЌЬиЛЏЮЊзЈгЊЪфЕМЙІФмЕФЕМЙмЗжзгЁЃ
ЁЁЁЁЕМЙмЗжзггыЙмАћЕФЧјБ№ЃЌжївЊдкгкЯИАћЕФЖЫБкдкЗЂг§Й§ГЬжаШмНтЃЌаЮГЩвЛИіЛђЪ§ИіДѓЕФПзЃЌГЦЮЊДЉПзЃЈperforationЃЉЃЌОпДЉПзЕФЖЫБкЬиГЦДЉПзАхЁЃдкФОжЪВПжаЃЌаэЖрЕМЙмЗжзгзнЯђЕиСЌНгГЩЯИАћааСаЃЌЭЈЙ§ДЉПзжБНгЙЕЭЈЃЌетбљЕФЕМЙмЗжзгСДОЭГЦЕМЙмЃЈvesselЃЉЃЈЭМ1Ѓ51ЃЌCЁЂDЁЂEЃЛЭМ1Ѓ52ЃЉЁЃЕМЙмГЄЖЬВЛвЛЃЌгЩМИРхУзЕНвЛУззѓгвЃЌгааЉЬйБОжВЮяПЩГЄДяЪ§УзЁЃЕМЙмЗжзгЕФЙмОЖвЛАувВБШЙмАћДжДѓЃЌвђДЫЃЌЕМЙмБШЙмАћОпгаНЯИпЕФЪфЫЎаЇТЪЁЃБЛзгжВЮяжаГ§СЫзюдЪМЕФРраЭЭтЃЌФОжЪВПжажївЊКЌгаЕМЙмЃЌЖјДѓЖрЪ§ТузгжВЮяКЭоЇРржВЮядђШБЗІЕМЙмЃЌетОЭЪЧБЛзгжВЮяИќФмЪЪгІТНЩњЛЗОГЕФживЊдвђжЎвЛЁЃ
ЁЁЁЁФОжЪВПжаЕФЯЫЮЌГЦЮЊФОЯЫЮЌЃЌЪЧФЉЖЫМтШёЕФЩьГЄЯИАћЃЌдкЭЌвЛжВЮяжаЃЌвЛАуБШЙмАћгаНЯКёЕФБкЃЌЖјЧвЧПСвФОжЪЛЏЃЌГЩЪьЪБдЩњжЪЬхЭЈГЃЫРЭіЃЌЕЋвВгааЉжВЮяЕФФОЯЫЮЌФмЩњЛюНЯГЄЕФЪБМфЁЃФОЯЫЮЌЕФДцдкЪЙФОжЪВПМцгажЇГжЕФЙІФмЁЃ
ЁЁЁЁФОжЪВПжаЩњЛюЕФБЁБкЯИАћЃЌГЦФОБЁБкЯИАћЃЌЫќУЧдкЗЂг§КѓЦкЃЌЯИАћБкЭЈГЃвВФОжЪЛЏЃЌетаЉЯИАћГЃКЌгаЕэЗлКЭНсОЇЃЌОпгаДЂВиЕФЙІФмЁЃ
ЁЁЁЁЂкШЭЦЄВП ШЭЦЄВПвВЪЧвЛжжИДКЯзщжЏЃЌАќКЌЩИЙмЗжзгЛђЩИАћЁЂАщАћЁЂБЁБкЯИАћЁЂЯЫЮЌЕШВЛЭЌРраЭЕФЯИАћЃЌЦфжагыгаЛњЮяЕФдЫЪфжБНггаЙиЕФЪЧЩИЙмЗжзгЛђЩИАћЁЃ
ЁЁЁЁ ЩИЙмЗжзгЃЈsieve-tube elementЛђsieve-tube memberЃЉгыЕМЙмЗжзгЯрЫЦЃЌЪЧЙмзДЯИАћЃЌдкжВЮяЬхжазнЯђСЌНгЃЌаЮГЩГЄЕФЯИАћааСаЃЌГЦЮЊЩИЙмЃЈsieve tubeЃЉЃЌЫќЪЧБЛзгжВЮяжаГЄОиРыдЫЪфЙтКЯВњЮяЕФНсЙЙЁЃ

 ЁЁ
ЁЁ
ЁЁЁЁЩИЙмЗжзгжЛОпГѕЩњБкЁЃБкЕФжївЊГЩЗжЪЧЙћНККЭЯЫЮЌЫиЁЃдкЫќЕФЩЯЯТЖЫБкЩЯЗжЛЏГіаэЖрНЯДѓЕФПзЃЌГЦЩИПзЃЈsieve poreЃЉЃЌОпЩИПзЕФЖЫБкЬиГЦЩИАхЃЈsieve plateЃЉЁЃДжЕФдЩњжЪСЊТчЫїЃЈconnecting strandЃЉДЉЙ§ЩИПзЪЙЩЯЯТСкНгЕФЩИЙмЗжзгЕФдЩњжЪЬхУмЧаЯрСЌЃЈЭМ1Ѓ53ЃЛЭМ1—54ЃЉЃЌдкИїСЊТчЫїЕФжмЮЇгаыныежЪЃЈcalloseЃЉЧЪАќЮЇЃЈЭМ1-54ЃЌBЃЉЁЃыныежЪЪєЬЧРрЃЌЪЧвЛжжβ-1ЃЌ3-ЦЯОлЬЧЁЃЩИЙмЗжзгЕФВрБкОпаэЖрЬиЛЏЕФГѕЩњЮЦПзГЁЃЌГЦЮЊЩИгђЃЈsive areaЃЉЃЌЦфЩЯЕФПзНЯвЛАуБЁБкЯИАћБкЩЯГѕЩњЮЦПзГЁЕФПзДѓЃЌБШАћМфСЌЫПИќДжЕФдЩњжЪЫПдкДЫЭЈЙ§ЃЌетЪЙЩИЙмЗжзггыВрСкЕФЯИАћгаИќУмЧаЕФЮяжЪНЛСїЁЃ
ЁЁЁЁЩИЙмЗжзгОпгаЩњЛюЕФдЩњжЪЬхЃЌЕЋЯИАћКЫдкЗЂг§Й§ГЬжазюКѓНтЬхЃЌвКХнФЄвВНтЬхЃЌЯИАћжЪжаБЃСєгаЯпСЃЬхЁЂжЪЬхЁЂP-ЕААзЬхКЭвЛВПЗжФкжЪЭјЁЃP-ЕААзЬхЪЧДѓВПЗжБЛзгжВЮяЕФЩИЙмЗжзгжаЬигаЕФНсЙЙЁЃдкВЛЭЌЕФжВЮяжжРржааЮзДВЛЭЌЃЌгаЙмзДЁЂЯЫЫПзДЁЂПХСЃзДКЭНсОЇаЮЕШЁЃдкЭЈГЃЧщПіЯТЃЌЫќУЧЗжЩЂдкЯИАћжЪжаЃЌЕЋЕБШЭЦЄВПЪмИЩШХЪБЃЌЫќУЧЛсОлМЏдкЩИПзДІаЮГЩеГвКШћЁЃЖдP-ЕААзЬхЕФЙІФмФПЧАЛЙВЛЧхГўЃЌгаШЫШЯЮЊЫќЪЧвЛжжЪеЫѕЕААзЃЌПЩФмдкЩИЙмдЫЪфгаЛњЮяжаЦ№зїгУЁЃЩИЙмжЪЬхдкГЩЗжКЭаЮзДЩЯгыЦфЫћЯИАћжаЕФВЛЭЌЃЌЫќУЧгаЕФКЌЕэЗлЃЌгаЕФКЌЕААзжЪЃЌдкЕчзгЯдЮЂОЕжаПЩЪЖБ№ГіЖржжаЮЬЌЃЌгаШЫвбАбетжжаЮЬЌЩЯЕФЧјБ№зїЮЊжВЮяЗжРржаЛЎЗжНЯДѓЗжРрШКЕФЬиеїЁЃ
ЁЁЁЁЩИЙмЗжзгЕФВрУцЭЈГЃгывЛИіЛђвЛСаАщАћЃЈcompanioncellЃЉЯрХўСкЃЌАщАћЪЧгыЩИЙмЗжзгЦ№дДгкЭЌвЛИідЪМЯИАћЕФБЁБкЯИАћЃЌОпгаЯИАћКЫМАИїРрЯИАћЦїЃЌгыЩИЙмЗжзгЯрСкЕФБкЩЯгаГэУмЕФЩИгђЃЌЗДгГГіЖўепЙиЯЕУмЧаЃЈЭМ1-54ЃЌAЃЉЁЃЯжСЫНтЃЌЩИЙмЕФдЫЪфЙІФмгыАщАћЕФДњаЛНєУмЯрЙиЁЃгаЕФжВЮяАщАћЗЂг§ЮЊДЋЕнЯИАћЁЃ

ЁЁ
ЁЁЁЁЩИЙмдЫЫЭбјЗжЕФЫйЖШУПаЁЪБПЩДя 10—100cmЁЃЭЈГЃЃЌЩИЙмЙІФмжЛгавЛИіЩњГЄМОЃЌЩйЪ§жВЮяПЩИќГЄЃЌШчЦЯЬбЁЂщВЁЂМюХюЕФЩИЙмПЩБЃГжЖўжСЖрФъЁЃдкЫЅРЯЛђанУпЕФЩИЙмжаЃЌдкЩИАхЩЯЛсДѓСПЛ§РлыныежЪЃЌаЮГЩЕцзДЕФыныеЬхЃЈcallusЃЉЗтБеЩИПзЃЌЕБДЮФъДКМОЩИЙмжиаТЛюЖЏЪБЃЌыныеЬхФмЯћЪЇЃЌСЊТчЫїгжФмжиаТЙЕЭЈЁЃДЫЭтЃЌЕБжВЮяЪмЕНЫ№ЩЫЕШЭтНчДЬМЄЪБЃЌЩИЙмЗжзгвВФмбИЫйаЮГЩыныежЪЃЌЗтБеЩИПзЃЌзшжЙгЊбјЮяЕФСїЪЇЁЃ
ЁЁЁЁТузгжВЮяКЭоЇРржВЮяжаЃЌвЛАуУЛгаЩИЙмЃЌдЫЪфгаЛњЮяЕФЗжзгЪЧЩИАћЃЈsieve cellЃЉЁЃЫќгыЩИЙмЗжзгЕФжївЊЧјБ№ЃЌдкгкЩИАћЕФЯИАћБкЩЯжЛгаЩИгђЃЈЭМ1-50ЃЌEЃЉЃЌдЩњжЪЬхжавВУЛгаP-ЕААзЬхЁЃ
ЁЁЁЁШЭЦЄВПЕФЯЫЮЌЦ№жЇГжзїгУЃЌШЭЦЄЯЫЮЌЕФЯИАћБкФОжЪЛЏГЬЖШНЯШѕЃЌЛђВЛФОжЪЛЏЃЌвђЖјжЪЕиНЯМсШЭЃЌгаНЯЧПЕФПЙЧњФгЕФФмСІЁЃаэЖржВЮяЕФШЭЦЄЯЫЮЌЗЂДяЃЌЯИАћГЄЁЂЯЫЮЌЫиКЌСПИпЁЂжЪЕиШсШэЃЌГЩЮЊЩЬгУЯЫЮЌЕФживЊРДдДЁЃР§ШчмбТщЁЂбЧТщЁЂТоВМТщЕШЕФШЭЦЄЯЫЮЌГЄЖјВЛФОжЪЛЏЃЌПЩзївТзХКЭеЪХёЕФдСЯЃЛЛЦТщЁЂбѓТщЁЂммТщЕШЕФШЭЦЄЯЫЮЌНЯЖЬЃЌгавЛЖЈГЬЖШЕФФОжЪЛЏЃЌПЩгУгкжЦТщДќКЭЩўЫїЕШЁЃ
ЁЁЁЁШЭЦЄВПЕФБЁБкЯИАћЃЌжївЊЦ№ДЂВиКЭКсЯђдЫЪфЕФзїгУЃЌГЃКЌгаНсОЇКЭИїРрДЂВиЮяЁЃ
ЁЁЁЁзлЩЯЫљЪіЃЌПЩвдСЫНтФОжЪВПКЭШЭЦЄВПЪЧжВЮяЬхжаЦ№ЪфЕМзїгУЕФЖўРрИДКЯзщжЏЃЌЫќУЧЕФзщГЩжаЗжБ№вдОпгаЪфЕМЙІФмЕФЙмзДЗжзг——ЕМЙмЗжзгЁЂЙмАћКЭЩИЙмЗжзгЛђЩИАћЮЊжїЃЌЫљвдЃЌдкаЮЬЌбЇЩЯЃЌгжНЋЖўепЗжБ№ЛђКЯГЦЮЊЮЌЙмзщжЏЁЃ
ЁЁЁЁЃЈ5ЃЉЗжУкНсЙЙ ФГаЉжВЮяЯИАћФмКЯГЩвЛаЉЬиЪтЕФгаЛњЮяЛђЮоЛњЮяЃЌВЂАбЫќУЧХХГіЬхЭтЁЂЯИАћЭтЛђЛ§РлгкЯИАћФкЃЌетжжЯжЯѓГЦЮЊЗжУкЯжЯѓЁЃжВЮяЗжУкЮяЕФжжРрЗБЖрЃЌгаЬЧРрЁЂЛгЗЂгЭЁЂгаЛњЫсЁЂЩњЮяМюЁЂЕЄФўЁЂЪїжЌЁЂгЭРрЁЂЕААзжЪЁЂУИЁЂЩБОњЫиЁЂЩњГЄЫиЁЂЮЌЩњЫиМАЖржжЮоЛњбЮЕШЃЌетаЉЗжУкЮядкжВЮяЕФЩњЛюжаЦ№зХЖржжзїгУЁЃР§ШчЃЌИљЕФЯИАћЗжУкгаЛњЫсЁЂЩњГЄЫиЁЂУИЕШЕНЭСШРжаЃЌЪЙФбШмадЕФбЮРрзЊЛЏГЩПЩШмадЕФЮяжЪЃЌФмБЛжВЮяЮќЪеРћгУЃЌЭЌЪБЃЌгжФмЮќв§вЛЖЈЕФЮЂЩњЮяЃЌЙЙГЩЬиЪтЕФИљМЪЮЂЩњЮяШКЃЌЮЊжВЮяНЁзГЩњГЄДДдьИќКУЕФЬѕМўЃЛжВЮяЗжУкУлжКЭЗМЯугЭЃЌФмв§геРЅГцЧАРДВЩУлЃЌАяжњДЋЗлЁЃФГаЉжВЮяЗжУкЮяФмвжжЦЛђЩБЫРВЁОњМАЦфЫћжВЮяЃЌЛђФмЖдЖЏЮяКЭШЫаЮГЩЖОКІЃЌгаРћгкБЃЛЄздЩэЁЃСэвЛаЉЗжУкЮяФмДйНјЦфЫћжВЮяЕФЩњГЄЃЌаЮГЩгавцЕФЯрЛЅвРДцЙиЯЕЕШЁЃвВгааЉЗжУкЮяЪЧжВЮяЕФХХаЙЮяЛђДЂВиЮяЁЃаэЖржВЮяЕФЗжУкЮяОпгаживЊЕФОМУМлжЕЃЌР§ШчЯ№НКЁЂЩњЦсЁЂЗМЯугЭЁЂУлжЕШЁЃ
ЁЁЁЁжВЮяВњЩњЗжУкЮяЕФЯИАћРДдДИївьЃЌаЮЬЌЖрбљЃЌЗжВМЗНЪНвВВЛОЁЯрЭЌЃЌгаЕФЕЅИіЗжЩЂгкЦфЫћзщжЏжаЃЌвВгаЕФМЏжаЗжВМЃЌЛђЬиЛЏГЩвЛЖЈНсЙЙЃЌЭГГЦЮЊЗжУкНсЙЙЁЃИљОнЗжУкЮяЪЧЗёХХГіЬхЭтЃЌЗжУкНсЙЙПЩЗжГЩЭтВПЕФЗжУкНсЙЙКЭФкВПЕФЗжУкНсЙЙСНДѓРрЁЃ
ЁЁЁЁЂйЭтВПЕФЗжУкНсЙЙЭтВПЕФЗжУкНсЙЙЦеБщЕФЬиеїЃЌЪЧЫќУЧЕФЯИАћФмЗжУкЮяжЪЕНжВЮяЬхЕФБэУцЁЃГЃМћЕФРраЭгаЯйБэЦЄЃЈglandular epidermisЃЉЁЂЯйУЋЃЈglandular hairЃЉЁЂУлЯйЃЈnectaryЃЉКЭХХЫЎЦїЃЈhydathodeЃЉЕШЁЃ
ЁЁЁЁЯйБэЦЄ МДжВЮяЬхФГаЉВПЮЛЕФБэЦЄЯИАћЮЊЯйадЃЌОпгаЗжУкЕФЙІФмЁЃР§ШчАЋЧЃХЃЃЈPetunia hybridaЃЉЁЂЦсЪїЃЈRhus vernicifluaЃЉЕШаэЖржВЮяЛЈЕФжљЭЗБэЦЄМДЪЧЯйБэЦЄЃЌЯИАћГЩШщЭЗзДЭЛЦ№ЁЂОпгаХЈКёЕФЯИАћжЪЃЌБЛгаБЁЕФНЧжЪВуЃЌФмЗжУкГіКЌгаЬЧЁЂАБЛљЫсЁЂЗгРрЛЏКЯЮяЕШзщГЩЕФжљЭЗвКЃЌРћгкеГзХЛЈЗлКЭПижЦЛЈЗлУШЗЂЁЃ

ЁЁ
ЁЁЁЁЯйУЋЯйУЋЪЧИїжжИДдгГЬЖШВЛЭЌЕФЁЂОпгаЗжУкЙІФмЕФБэЦЄУЋзДИНЪєЮяЃЈЭМ1-55ЃЉЁЃЯйУЋвЛАуОпгаЭЗВПКЭБњВПЖўВПЗжЃЌЭЗВПгЩЕЅИіЛђЖрИіВњЩњЗжУкЮяЕФЯИАћзщГЩЁЃБњВПЪЧгЩВЛОпЗжУкЙІФмЕФБЁБкЯИАћзщГЩЃЌзХЩњгкБэЦЄЩЯЁЃбЌвТВнЃЈLavandulaangusti-foliaЃЉЁЂУоЁЂбЬВнЁЂЬьѓУПћЁЂБЁКЩЕШжВЮяЕФОЅКЭвЖЩЯЕФЯйУЋОљЪЧШчДЫЁЃнЁТщЪєЃЈUrticaЃЉЕФѓЇУЋОпгаЬиЪтЕФНсЙЙЃЌЫќЪЧЕЅИіЕФЗжУкЯИАћЃЌЫЦвЛИіЛљВПХђДѓЕФУЋЯИЙмЃЌЖЅВПЗтБеЮЊаЁдВЧђзДЁЃЕБУЋгыЦЄЗєНгДЅЪБЃЌдВЧђЖЅВПдгаЕФЗьЯпЦЦСбЃЌТЖГіЗцРћЕФБпдЕЃЌДЬНјЦЄЗєЃЌдйгЩХнзДЛљВПНЋКЌгаЕФвЯЫсКЭзщжЏАЗЕШЕФвКЬхМЗНјЩЫПкЁЃаэЖрФОБОжВЮяШчРцЪєЃЈPyrusЃЉЁЂЩНКЫЬвЪєЃЈCaryaЃЉЁЂшыФОЪєЃЈBe-tuaЃЉЕШЃЌдкгзаЁЕФвЖЦЌЩЯОпгаеГвКУЋЃЌЗжУкЪїНКРрЮяжЪИВИЧећИівЖбПЃЌЗТЗ№ИјбПЬсЙЉСЫвЛИіБЃЛЄадЭтЬзЁЃЪГГцжВЮяЕФБфЬЌвЖЩЯЃЌПЩвдгаЖржжЯйУЋЗжБ№ЗжУкУлТЖЁЂеГвККЭЯћЛЏУИЕШЃЌгав§геЁЂеГзХКЭЯћЛЏРЅГцЕФзїгУЁЃ
ЁЁЁЁ ХХЫЎЦї ХХЫЎЦїЪЧжВЮяНЋЬхФкЙ§ЪЃЕФЫЎЗжХХГіЕНЬхБэЕФНсЙЙЁЃЫќЕФХХЫЎЙ§ГЬГЦЮЊЭТЫЎЃЈguttationЃЉЁЃХХЫЎЦїгЩЫЎПзЁЂЭЈЫЎзщжЏКЭЮЌЙмЪјзщГЩЃЈЭМ1-56ЃЉЃЌЫЎПзЃЈwaterPoreЃЉДѓЖрДцдкгквЖМтЛђвЖТЬЃЌЫќУЧЪЧвЛаЉБфЬЌЕФЦјПзЃЌБЃЮРЯИАћвбЪЇШЅСЫЙиБеПзЕФФмСІЁЃЭЈЫЎзщжЏЃЈepithemЃЉЪЧЫЎПзЯТЕФвЛЭХБфЬЌвЖШтзщжЏЃЌЯИАћХХСаЪшЫЩЃЌЮовЖТЬЬхЁЃЕБжВЮяЬхФкЫЎЗжЖргрЪБЃЌЫЎЭЈЙ§аЁвЖТіФЉЖЫЕФЙмАћЃЌСїОЭЈЫЎзщжЏЕФЯИАћМфЯЖЃЌзюжеДгЫЎПзХХГіЬхЭтЃЌаЮГЩЭТЫЎЁЃаэЖржВЮяЃЌШчКЕН№СЋЁЂОэаФВЫЁЂЗЌЧбЁЂВннЎЁЂЕигмЕШЖМгаУїЯдЕФЭТЫЎЯжЯѓЃЌИЁвЖЫЎЩњжВЮяЁЂШчСтЁЂЫЏСЋЕШЭТЫЎИќЮЊЦеБщЁЃ
ЁЁЁЁУлЯй УлЯйЪЧвЛжжЗжУкЬЧвКЕФЭтВПЗжУкНсЙЙЃЌДцдкгкаэЖрГцУНЛЈжВЮяЕФЛЈВПЁЃЗжУкЛЈУлЃЌЬсЙЉДЋЗлРЅГцЫљашЕФЪГЮяЃЌгыЛЈЕФЩЋВЪКЭЯуЮЖЯрХфКЯЃЌЪЪгІГцУНДЋЗлЕФЬиеїЃЌетРрУлЯйГЦЛЈУлЯйЁЃдквЛаЉжВЮягЊбјЬхЕФЕиЩЯВПЗжЃЌШчОЅЁЂвЖЁЂвЖБњКЭАњЦЌЕШВПЮЛвВДцдкУлЯйЃЌетаЉУлЯйГЦЛЈЭтУлЯйЃЌЫќУЧБЛШЯЮЊЪЧдкжВЮяНјЛЏЙ§ГЬжагыеав§ТьвЯвдБмУтЦфЫћЪГВнКІГцЕФЮЃКІгаЙиЁЃЛЈЭтУлЯйВЛНіДцдкгкБЛзгжВЮяЃЌдкФГаЉоЇРржВЮяЕФвЖЩЯвВгаДцдкЁЃУлЯйЕФаЮЬЌЖрбљЃЌгаЕФЮоЬиЪтЭтаЮЃЌжЛЪЧЯйБэЦЄРраЭЃЌШчзЯдЦгЂЕФЛЈУлЯйЪЧдкалШяКЭДЦШяжЎМфЕФЛЈЭаБэЦЄОпЯйадЃЌФмЗжУкЛЈУлЃЛКЕН№СЋЪЧЛЈОрЕФФкБэЦЄФмЗжУкЛЈУлЁЃгаЕФжВЮяУлЯйЗжЛЏГЩОпвЛЖЈЭтаЮЕФЬиЪтНсЙЙЃЌШчгЭВЫЛЈУлЯйдкЛЈЭаЩЯГЩ4ИіТЬЩЋЕФаЁПХСЃЃЛШ§ЩЋнРЕФЛЈУлЯйдкЖўИіалШяЩЯЃЌЪЧвЉИєбгЩьГЩЕФЖўИіАєзДЮяЩьШыЛЈОрФкЃЛЮкшъКЭвЛЦЗКьЕФЛЈЭтУлЯйЗжБ№ГЩХЬзДКЭБзДДцдкгквЖЭЉКЭЛЈађзмАњЦЌЩЯЁЃУлЯйЕФФкВПНсЙЙБШНЯвЛжТЃЌЗжУкзщжЏДѓЖрАќРЈБэЦЄМАБэЦЄЯТМИВуБЁБкЯИАћЁЃетаЉЯИАћЬхЛ§НЯаЁЃЌЯИАћжЪХЈЁЂКЫНЯДѓЃЌГЃОпгаЗЂДяЕФФкжЪЭјКЭИпЖћЛљЬхЃЌгаЪБЗЂг§ГЩДЋЕнЯИАћЁЃППНќЗжУкзщжЏГЃОпгаЮЌЙмЪјЁЃгЩгкУлЕФдСЯРДздШЭЦЄВПЕФжвКЃЌвђДЫЃЌетаЉЮЌЙмЪјжаКЌгаЕФШЭЦЄВПКЭФОжЪВПЕФБШР§гыУлжЕФГЩЗжгаЙиЃЌЕБШЭЦЄВПЗЂДяЪБЃЌУлжаЬЧЗжКЌСПНЯИпЃЌЗДжЎЃЌФОжЪВПЗЂДяЪБЃЌЬЧЗжКЌСПНЕЕЭЃЌЫЎЗжКЌСПдіИпЁЃ

ЁЁ
ЁЁЁЁЂкФкВПЕФЗжУкНсЙЙЗжУкЮяВЛХХЕНЬхЭтЕФЗжУкНсЙЙЃЌГЦЮЊФкВПЕФЗжУкНсЙЙЃЌАќРЈЗжУкЯИАћЃЈsecretorycellЃЉЁЂЗжУкЧЛЃЈsecretorycavityЃЉЛђЗжУкЕРЃЈsecretorycanalЃЉвдМАШщжЙмЃЈlaticiferЃЉЁЃ
ЁЁЁЁЗжУкЯИАћ ЗжУкЯИАћПЩвдЪЧЩњЛюЯИАћЛђЗЧЩњЛюЯИАћЃЌЕЋдкЯИАћЧЛФкЖМЛ§ОлгаЬиЪтЕФЗжУкЮяЁЃЫќУЧвЛАуЮЊБЁБкЯИАћЃЌЕЅИіЕиЗжЩЂгкЦфЫћЯИАћжЎжаЃЌЯИАћЬхЛ§ЭЈГЃУїЯдЕиНЯжмЮЇЯИАћЮЊДѓЃЌгШЦфдкГЄЖШЩЯИќЮЊЯджјЃЌвђДЫШнвзЪЖБ№ЁЃИљОнЗжУкЮяжЪЕФРраЭЃЌПЩЗжЮЊгЭЯИАћЃЈеСПЦЁЂФОРМПЦЁЂРАУЗПЦЕШЃЉЁЂеГвКЯИАћЃЈЯЩШЫеЦПЦЁЂНѕПћПЦЁЂщВПЦЕШЃЉЁЂКЌОЇЯИАћЃЈЩЃПЦЁЂЪЏЫтПЦЁЂбМѕХВнПЦЕШЃЉЁЂїЗжЪЯИАћЃЈЦЯЬбПЦЁЂОАЬьПЦЁЂЖЙПЦЁЂЧОоБПЦЕШЃЉвдМАНцзгУИЯИАћЃЈАзЛЈВЫПЦЁЂЪЎзжЛЈПЦЃЉЕШЁЃ
ЁЁЁЁЗжУкЧЛКЭЗжУкЕР ЫќУЧЪЧжВЮяЬхФкжќВиЗжУкЮяЕФЧЛЛђЙмЕРЁЃЫќУЧЛђЪЧвђВПЗжЯИАћНтЬхКѓаЮГЩЕФЃЈШмЩњЕФЃЌlysigenousЃЉЃЌЛђЪЧвђЯИАћжаВуШмНтЃЌЯИАћЯрЛЅЗжПЊЖјаЮГЩЕФЃЈСбЩњЕФЃЌschizo-genousЃЉЃЌЛђЪЧетЖўжжЗНЪНЯрНсКЯЖјаЮГЩЕФЃЈСбШмЩњЕФЃЌschizo-lysigenousЃЉЁЃР§ШчИЬщйвЖзгМАЙћЦЄжаЭЈГЃПДЕНЕФЛЦЩЋЭИУїаЁЕуЃЌБуЪЧШмЩњЗНЪНаЮГЩЕФЗжУкЧЛЃЌзюГѕЪЧВПЗжЯИАћжааЮГЩЗМЯугЭЃЌКѓРДетаЉЯИАћЦЦСбЃЌФкКЌЮяЪЭЗХЕНШмЩњЕФЧЛФкЁЃдкетжжШмЩњЧЛЕФжмЮЇПЩвдПДЕНгаВПЗжЫ№ЛЕЕФЯИАћЮЛгкЧЛЕФжмЮЇЃЈЭМ1-57ЃЉЁЃЫЩАиРрФОжЪВПжаЕФЪїжЌЕРКЭЦсЪїШЭЦЄВПжаЕФЦсжЕРЪЧСбЩњаЭЕФЗжУкЕРЃЌЫќУЧЪЧЗжУкЯИАћжЎМфЕФжаВуШмНтаЮГЩЕФзнЯђЛђКсЯђЕФГЄаЮЯИАћМфЯЖЃЌЭъећЕФЗжУкЯИАћГФдкЗжУкЕРЕФжмЮЇЃЌЪїжЌЛђЦсвКгЩетаЉЯИАћХХГіЃЌЛ§РлдкЙмЕРжаЃЈЭМ1-58ЃЉЁЃЙћЪєЃЈMangiferaЃЉЕФвЖКЭОЅжаЕФЗжУкЕРЪЧСбШмЩњЦ№дДЕФЁЃ

ЁЁЁЁБОЮФБъЬтЃКЕквЛеТ жВЮяЯИАћКЭзщжЏ(10)
ЁЁЁЁЪжЛњвГУцЃКhttp://m.dljs.net/dlsk/zhiwuxue/59446.html
ЁЁЁЁБОЮФЕижЗЃКhttp://www.dljs.net/dlsk/zhiwuxue/59446.html
