��
��������
��
�����ɻ�����ɢ���ij��컨�ۣ�����һ����ý�������������͵�ͬһ������һ���Ĵ�����ͷ�ϵĹ��̣���Ϊ���ۣ�pollination����
����������������ֳ������ȱ�ٵĻ��ڣ�û�д��ۣ�Ҳ�Ͳ���������ܾ����á���Ϊ������ֳ�����еĴ�����——��ϸ�����Dz�����������ģ��������������ӷ����ڵ������Ҫ���ȫ��������ֳ���̣���һ������ʹ����������——��ϸ���Ļ���������ӽ������۾���������һ�����á�
������Σ����۵��ȷ���Ҫͨ����ͷ�������ʵĴ̼����ú�������������һ������£�����ֻ���䵽��ͷ�ϲ�����Ȼ���ȷ���������ͷ��ɢ��������IJ�ֲͬ�ﻨ�۾���ѡ�����ã�ֻ��һ���������͵Ļ��۲��ܵõ��ȷ����������Ļ��ۻ��ܵ���ͷ�����ƣ���������ֳ���á�����Ϊ������ֳ�����л��۵���Դ���죬�ܾ����������������ͬ���Ľ��Ҳ�Ͳ�ͬ��������һ������������Ӧ�Ե�ǿ����Ҳ�������ʵIJ�ͬ�����ֲ��졣���ֲ����Ŵ��ͱ����������й�ϵ�ġ�
������һ���Ի��������컨���ۼ�������ѧ����
������������һ�������ַ�ʽ��һ���Ի����ۣ�self-pollination������һ���컨���ۣ�cross pollination���������ִ��۷�ʽ����Ȼ�綼�ձ���ڡ�
 ����
����
����1���Ի����� ���۴ӻ�����ɢ�����䵽ͬһ������ͷ�ϵĴ�������Ϊ�Ի����ۡ�����ֲ���еĴ���С���㶹�����ѵȣ������������۵ġ���ʵ��Ӧ���ϣ��Ի����۵ĸ����ָ�ڹ���������ͬƷ�ּ�Ĵ��ۣ�����ũҵ��ͬ���컨��Ĵ��ۡ�
�����Ի�����ֲ��Ļ���Ȼ�ǣ���1�����Ի����������ﳣΧ�ƴ�����������Ұ��úܽ������Ի����������ڱ�������ͷ�ϣ���2������Ļ����Һʹ�������ұ�����ͬʱ����ģ���3���������ͷ���ڱ����Ļ����ȷ��ͻ��۹��������ӵķ���û���κ������谭��ͼ4��46����
�������۷�ʽ�еıջ����ۺͱջ��ܾ���cleistogamy����һ�ֵ��͵��Ի����ۣ�����һ��Ŀ������ۺͿ����ܾ���chasmogamy���Dz�ͬ�ġ�����ֲ��Ļ����������ſ������Ѿ�����ܾ����á����ǵĻ���ֱ���ڻ��������ȷ������۹ܴ��������ҵıڣ�����ͷ����������ܾ�����ˣ��ϸ�ؽ������ڴ�����һ���ڣ������㶹�����ӡ��仨���ȡ��ջ��ܾ�����Ȼ����һ�ֺ�������Ӧ����ֲ���ڻ������������ڿ�������ʱ���ջ��ܾ����ֲ�����һ���㣬�����ֳ���̣����һ��ۿ��Բ�������ˮ����ʪ���������ʳ��
����2.�컨����һ�仨�Ļ��۴��͵�ͬһֲ���ֲͬ����һ�仨����ͷ�ϵĴ��۷�ʽ�����컨���ۡ��ڹ��������ϣ���ͬƷ�ּ�Ĵ��ۺ����������ϲ�ֲͬ���Ĵ��ۣ�Ҳ���컨���ۡ�
�����컨���۵�ֲ��ͻ����ڽṹ�������ϲ�����һЩ�������Ӧ�Ա仯��ʹ�Ի����۳�Ϊ�����ܣ���Ҫ�����ڣ���1�������ԣ������Ǵ�������ֲ���2�����Ի���������ʹ��ﲻͬʱ���죬�ڴơ��������������У�����������ģ������ס�ݫ�ĵȣ����Ǵ�������ģ���ľ������˵ȣ�ͼ4��47������3���ơ������쳤����λ��Ҳ���ܽ����Ի����ۣ��籨������ͼ4��48������4���������ڱ�����ͷ�ϲ����ȷ���������ȫ�����Դﵽ�ܾ��Ľ�������������顢�ҡ��桢ƻ�������ѵȡ�

 ��
��
�����컨������ֲ���Ƚ��ձ�ش����ţ����Ի�������ȣ���һ�ֽ����ķ�ʽ���������ڵ��Ի����۶�ֲ�����к��ģ���ʹ�������������˥�ˣ�����ũҵ����ʵ�����ѵõ�֤��������С�����Ի����۵�ֲ�������������Ի����ۣ�30—40������˥�˶�ʧȥ�����ֵ��ͬ������������10—15����Ի����ۺ�Ҳ�����ͬ�������컨���ۺ������ܾ��Ͳ����������ǵĺ����������ǿ�������������Ӧ�ԡ�����ľ������ڵĹ۲��о���ָ����“�����Ի����۶�ֲ�ﱾ�����к��ģ����컨���۶�ֲ���������”�����Ľ�����ũҵʵ������ȫһ�µġ�
�����Ի����ۡ������ܾ�֮�����к������컨���ۡ������ܾ�֮�������棬����Ϊ�Ի�����ֲ�����������������ӣ��Ǵ���ͬһ���������£������ӵ��Ŵ���ȱ���ֻ����ã������С�������ںϺ�����ĺ������������Ӧ��С�����컨�������ڴơ����������ڱ˴˲���ȫ��ͬ�����������²����ģ��Ŵ��Ծ߽ϴ���죬�ںϺ�����ĺ����Ҳ���н�ǿ������������Ӧ�ԡ�
������Ȼ�Ի�������һ��ԭʼ�Ĵ�����ʽ���Ժ��Ҳ�������棬������Ȼ��ȴ��������������������Ϊ�Ի����۶�ijЩֲ����˵���������ġ����컨����ȱ������ķ硢���ý����������ʹ���۲��ܽ��е�ʱ���Ի������ֲ�����һȱ�㡣������������ָ��������ֲ����˵���������ܾ���������ֳ���ӣ��ܱȲ���ֳ���ӻ�ֳ���������ú�Щ���ο�����Ȼ��û��һ��ֲ���Ǿ����Ի����۵ģ��������м��ܻ��бȽ��ٵ�һ����ֲ�����ڽ����컨���ۡ����ԣ��������Ի����۵�ֲ�����࣬�����ձ���ڡ�
������������ý���ͳ�ý��
����ֲ������컨���ۣ������������������İ��������ܰѻ��۴���������������ͷ��ȥ�����ͻ��۵�ý���з��������桢���ˮ����Ϊ�ձ���Ƿ�����档���ֲ�ͬ�������۵Ļ�����������һЩ�������Ӧ�Խṹ��ʹ���۵õ���֤��
����1����ý�� ���������ͻ��۵Ĵ��۷�ʽ�Ʒ�ý��anemophily�����������ʽ���۵Ļ����Ʒ�ý����anemophilous flower�����ݹ��ƣ�Լ��1/10�ı���ֲ���Ƿ�ý�ģ��ֺ̱���ֲ���ľ��ֲ���е��ݡ����ľ�ȶ��Ƿ�ýֲ�
������ýֲ��Ļ����ܼ�����״�����|�軨��ȣ��ܲ����������ۣ�ͬʱɢ�ţ�����ͼ 4��28��C��������һ�����ᡢ�������⻬�����ױ��紵�͡��̱���ֲ����С��ˮ���ȵĻ�˿�ر�ϸ������ҩ���ھ��������Ƭ֮�⣬�ܷ����Ĵ�����ʹ�������۴�ɢ��������ȥ����ý���Ļ��������ϳ�����ͷ������״���߳����⣬���ӽ��ܻ��۵Ļ��ᡣ������ýֲ������Ҷ������ϰ�ԣ���������֦�ϵ�Ҷչ��֮ǰ��ɢ���Ļ����ܷ紵��ʱ�����Բ�����֦Ҷ���赲�����⣬��ýֲ��Ҳ���Ǵ����컨�����꣬��������ʧ��������ζ��ɫ����Щ�����DZ�Ҫ���������еķ�ý�����������Եģ�Ҳ��������̱���ֲ��Ļ������Եģ��㡢�ʵ�ֲ��Ļ�Ҳ������
����2����ý�� ������Ϊý����д��۷�ʽ�ijƳ�ý��entomophily�����������ʽ���۵Ļ����Ƴ�ý����entomophilous flower���������л�ֲ�����������洫�۵ģ������Ĵ��������з��ࡢ���ࡢ���ࡢӬ��ȣ���Щ���������ڻ���֮�䣬����Ϊ���ڻ��в��ѣ������Ի���Ϊ��Ϣ���������Dz�ʳ���ۡ�������Ϊʳ�ϡ�����Щ��У����ɱ����Ҫ�뻨�Ӵ�������Ҳ�ͽ����۴��ͳ�ȥ��
������Ӧ���洫�۵Ļ���һ���������������
������ý������������ζ���������档��ֲͬ��ɢ������ζ��ͬ��������������������Ҳ��һ������ϲ����ģ�Ҳ��ϲ����ġ�
������ý������ܲ���֭�����ٻ��Ƿֲ��ڻ��ĸ������֣����Ƿ�չ����������١����۾����ں�����ڻ��ĵײ������еľ��ڡ����۱�¶����ģ������ɼ׳桢Ӭ�Ͷ��ǵķ��ࡢ��������������������ڻ���֮�ڵģ���Ϊ���ǵĵ���Ͷ�������ȡ������ȡ��ʱ��������ճ�����������϶���������ȥ��
������ý������һ�ص��ǻ�������������и�������ɫ�ʡ�һ����俪�ŵĻ���졢�ơ��ϵ���ɫ�������俪�ŵĶി��ɫ��ֻ��ҹ���Ķ�����ʶ�𣬰������ۡ�
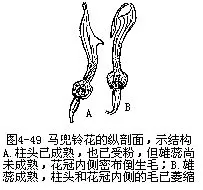
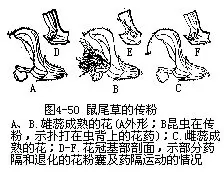
�������⣬��ý���ڽṹ��Ҳ���ʹ��۵�������γɻ�Ϊ��Ӧ�Ĺ�ϵ��������Ĵ�С�����Ρ��ṹ����Ϊ���뻨�Ĵ�С���ṹ�����ٵ�λ�õȣ�����������صġ����������廨������������Ϊ��Ͳ�����ơ��������죬����λ�ڻ�Ͳ���������⣬�ڻ�Ͳ�ڱ�����б������ĵ�ë����Щ��������Ĵ���������ء�������Ĵ����ǿ�һЩС����Ϊý��ģ������ڴ������ʱ��С��˳�ŵ�ë���뻨Ͳ�������ۣ���ʱ����Я���Ļ��۾ͱ����͵��������ͷ�ϡ���Ϊ��Ͳ�ڱڵĵ�ë��δ��ή��С��Ϊ��ë���ڻ��ڣ�һʱ��������ֱ����ҩ���죬����ɢ������ë����ή��Ϊ�����������ͨ�������������������Ҳ��ճ�ϴ������ۣ���������һ������ʱ���ְѻ��۴�����һ������ͷ��ȥ��δ�ڷ�ʱ������Ļ�����ֱ���ģ���������ɺ�����ͼ4��49����������β�ݵĴ��ۣ�Ҳ������Ȥζ�ġ���β�������ο�ֲ����Ļ��ࡢ���ڶ������ɹ�״����5Ƭ������ϲ�ȴ���ѳɴ��Σ���2Ƭ�ϳ�ͷ��״���ϴ�����3Ƭ���ϣ��γ��´�����ˮƽ����������ϴ���������2ö�����1������������ṹ���⣬��Ϊ1����ĸܸ�ϵͳ������ҩ���ӳ��ɸܸ˵ı����ϱ۳���������2������Ļ����ң��±۶̣������Ҳ������չΪ��Ƭ״��2����ı�Ƭ״�±�ͬλ�ڻ��ڹܵĺ�������ס���ڹܵ���ڡ����۷���뻨�ڹܵ������ʱ����Ҫͣ�����´��ϣ�Ȼ����ͷ���ƶ���Ƭ�����ܽ��뻨������ȡ���ۣ����ڸܸ˵ĵ���������Ƭ�����ƶ�ʱ���ϲ��ij�������������ʹ���˵Ļ�ҩ���䵽�۷�ı���������Ҳ��ɢ�������汳�ϡ�����������β�ݵĻ����϶̣������۳���ɢ���Ժ�����ʼ�쳤����ͷ���ôﵽ���汳����λ���ϣ��ȵ����л��۵���һ�۷������һ���ڲ���ʱ�����ϵĻ�������Ϳ�����µ���ͷ�ϣ���ɴ������ã�ͼ4��50����������˵����ý��������Ӧ�ṹ�����ӻ��ܶ࣬�Ͳ�һһ�����ˡ�
������ý���Ļ�����һ��ȷ�ý����Ҫ������ڴֲڣ����д�ͻ����ҩ�ѿ�ʱ��Ϊ�紵ɢ������ճ�ڻ�ҩ�ϣ������ڷû�����ʱ���״������������ܣ��������ͷҲ����ճҺ���ڣ�����һ���Ӵ�������ճס����������ҲԶ�Ϸ�ýΪ�١�
 ��
��
����3���������۷�ʽ ����ý�ͳ�ý�����⣬ˮ������ֲ���еĽ����塢���塢ˮ��ȶ��ǽ�ˮ�������۵ģ����ഫ�۷�ʽ��ˮý��hydrophily������������ֲ���Ǵ�������ģ�����������ˮ�ף����ۻ�����ʱ�������ۻ��Ի������䣬����ˮ�濪�ţ�ͬʱ�ƻ�����Ѹ���ӳ����Ѵƻ�����ˮ�棬���ۻ�Ʈ���ƻ�ʱ�����ֻ���ˮ����������ͷ���ۻ���ҩ�Ӵ�����ɴ��ۺ��ܾ����̣��Ժ�ƻ��Ļ������¾���������״���Ѵ������ˮ�ף���һ�������ɹ�ʵ�����ӣ�ͼ4��51����
��������������ഫ�۵ij���ý��ornithophily�������۵���һЩС�εķ���Heliothrix au-rita����ͷ���г������ȡ����ʱ�ѻ��۴�����ͼ4��52������ţ�������С����Ҳ�ܴ��ۣ�����������

 ��
��
�����������˹������ڷ�
�����컨�������������ܵ��������������ƣ��ò������۵Ļ��ᣬ���ý����û�з磬��ý������������µͣ���ȱ���㹻����ɳ�����۵ȣ��Ӷ����ʹ��ۺ��ܾ��Ļ��ᣬӰ�쵽��ʵ�����ӵIJ�������ũҵ�����ϳ������˹������ڷ۵ķ������Կ˷������������ʹ���۵ò�����֤��ȱ�ݣ��ԴﵽԤ�ڵIJ�������Ʒ�ָ�׳�Ĺ����У�Ҳ��Ҫ��ȡ�˹������ڷۣ��ԴﵽԤ�ڵ�Ŀ�ġ��˹������ڷۿ��Դ���������ͷ�ϵĻ�������ʹ�����������ļ�����������������ӣ�ø�ķ�ӦҲ��Ӧ���˼�ǿ���ٽ������ȷ��ͻ��۹����������ã��ܾ��ʿ��Եõ��ܴ���ߡ���������һ�����������£������������죬���������ʱ�ѵò�����ʱ�Ĵ��ۣ�������붥�������γ�ȱ���������˲������˹������ڷ۾��ܿ˷���һȱ�㣬ʹ�������8—10%���ҡ��������տ�����Ȼ���������£��ձ����϶࣬��������˹������ڷۣ�ͬ������߽�ʵ�ʺͺ�������
�����˹������ڷ۵ľ��巽�����ڲ�ͬ���ﲻ��ȫһ����һ�����ȴ������ϲɼ����ۣ�Ȼ������������ͷ�ϣ����߽��ռ��Ļ��ۣ��ڵ��º���������¼��Ա�أ������Ժ����á�
��
�����ܾ�
��
��������������ɺ��۱�����ͷ���ȷ��ɻ��۹ܣ����ڲ�������������——���ӣ�ͨ�����۹ܵ��쳤��ֱ������������ڲ�������ϸ���ͼ��˻����ںϡ������������ӻ����ںϵĹ��̣����ܾ����ã�fertilization������������ֳ���̵���Ҫ�Ρ��ܾ���������һ������Ϊ���ӡ�
������һ������������ͷ�ϵ��ȷ�
����������ͷ�ϵĻ�����������ͷ���ڵ�ճҺ��ճס���Ժ۵��ڱ����ȷ��״�����ͻ�����������쳤���γɻ��۹ܣ���һ���̣��ƻ��������ȷ�����ʹ���������ȷ��������ɻ��۹ܵ������Ƕ��ģ�������ͷ�ķ�����ͻ��۱��������ø�ʹ�л���ͷ���ڵ�ճ�����ʣ����Դ�ʹ�����ȷ�������ֹ�������ڸ����������ճ�����ʵ���Ҫ�ɷ���ˮ�����ࡢ���ܲ��ء�����ø��ά���صȡ����ڷ��������ɳɷ���ֲ��������죬�����������ͷ�ϵĸ���ֲ�ﻨ�۲�����Ӱ��Ҳ�Ͳ�ͬ��
��������������ͷ�Ϻ���������Χ��ȡˮ�֣���ˮ��Ļ�������������Ѹ����ǿ���ۺ������������࣬�����ʵĺϳ�Ҳ����������ߡ�ͬʱ������������ˮ������������߶�����Ҳ��ǿ��Ծ�������ܶ����С�ݣ����Ŷ���ø��������ʣ�ѭ��ϸ������ǰ�����ķ����ͷų�С�ݣ����뻨�۹ܱڵĽ��ɡ���ˮ�Ļ�����Ӫ��ϸ����Һ�ݻ���ǿ��ϸ���ڲ��������࣬ϸ������ѹ���ӣ������ʹ���������ڱ�����һ���������ȷ���ͻ�����γɻ��۹ܣ�pollen tube����ͼ4��53����

��
������ֲͬ���ͷ�ķ������ڳɷֺ�Ũ���ϳ�������ͬ���ر��Ƿ������ʵı仯���Ի����ȷ������ٽ������Ƶ�“ѡ��”���á����⣬ʵ��֤����������ӶԻ��۹����������ã�����Լ��ٻ������ѣ�����۵��ȷ��ʣ�����ʹ���۹������������յ����۹ܲ���һ���������ԡ��е�ֲ�����ʱ��ͷ�ϵ�ë״���ͷų����ַ��㻯��������յ������ȷ������á��������������ø���ִ�л�����ǻ������ȷ����������أ��������ڻ��۱ڵĶ���ˮ��ø������ˮ�Ӵ����ɱ����˳����Ի����ȷ��ͻ��۹ܳ�����ͷ������Ҫ���á����绨�����ͻ��۹��еĽ���ø����ʹ��ͷ������ͷ״ͻ��Ľ����ܽ⣬Ϊ���۹ܵ���������ͨ���������ڵĴ�л���ʣ���Ϊ���۹ܵ���������ṩ���ʻ�����
�����䵽��ͷ�ϵĻ�����Ȼ�ܶ࣬������ȫ�����ȷ��ģ��κ�һ��ֲ�↑��ʱ���Խ��ܱ���ֲ��Ļ��ۣ�ͬʱҲ���ܽ��ܲ���ͬ��ֲ��Ļ��ۡ�������ͬ�ֵģ����ڣ����Dz�ͬ�ֵģ��ּ䣩��ֻ�н�������ױ����Ŵ����Ͻ�Ϊ�ӽ�������Ȳ�����Ҳ����С�����п���ʵ�����ԵĽ��䣬�����˵�������ֲ��㷺�ر���Ϊͬһ���ڵ��컨�ܾ��ǿ��͵ģ������Ŵ��ϲ����ش������¾Ͳ����͡����͵Ļ�������ͷ�ϻ��Dz����ȷ��������ȷ��۹��������������ܴ�����ͷ�����ǻ��۹��ڻ����ڵ������ܵ����ƣ����ܴﵽ�ӷ������Դӻ����䵽��ͷ�Ϻ���ͷ�Ի��۾ͽ���“ʶ��”��“ѡ��”�����͵Ļ�������“�Ͽ�”�����͵ľ�����“�ܾ�”��
��������ܾ��ͻ��͵�������ʲô���ݽ�������ϸ������ѧ�ĽǶ��о���֤�������������֯���ʶ��Ӧ�Ǿ����ڻ��۱��Ϻͱ��ڵĵ����ʺ���ͷ���浰���ʱ�Ĥ֮������ϵ���⼸�ֵ�������ʶ���ͻ��͵����ʻ��������컨������Χ�ĵ���������ڵ����ڱڵ������֣�ǰ���ǻ����ڳ���ʱ�ɻ����ұڵ���ձ��ϸ�������ڲ��������ڻ�����ڵ�ǻ϶�ڣ������ǻ����ڳ���ʱ�ɻ���ϸ�����������������ڻ����ڱ��ϵģ����ȷ��������Ϊ�ḻ�����⣬����ͷ���渲��һ����ˮ�ĵ����ʱ�Ĥ�����ճ��Ĥ������ճס������������“ʶ��”���۵�������������������ͷ���ܵ���ʪʱ����ڵ����ڼ������ڱ��ͷų��������ڱڵ���ͨ���ȷ���Ҳ�ӻ����ݳ��������ֻ��۵�����ڻ��۸�������ͷ���棬����ͷ����ĵ��������á��͵Ļ����ܴ���ͷ��ˮ����ʼ�ȷ������͵Ļ����“�ܾ�”����ʱ�ڻ����ȷ����ڲſ�ʼ����Ļ��۹ܶ��γ�������������ֹ�˻��۹ܵ��������е�������ͷ�������ͻϸ���ڲ������������������������۹ܵĴ��롣
�������Խ����ӽ������������ܾ��IJ����ԣ����²��У������ֹ���������ѣ����Խ������ڿ˷������Ե��ϰ�����������о������ж��ִ�ʩ���Բ��ã��磺�û�ϻ����ڷۣ��������ڷۣ��ڷ�ǰ�س���ͷ��ض̻������ӷ����ڷۻ��Թ��ܾ��ȡ�
������������ͷ���������ȷ��ģ������ס��ݵȣ�������Ҫ������������������һЩʱ�����ȷ��ģ�������С����˵ȡ�����ʪ�ȹ��ߣ������¹��ͣ����ܴﵽ�ȷ�����Ҫ��ʪ�Ȼ��¶�ʱ���ȷ��ͻ��ܵ�Ӱ�졣����ʱ���������������������Ž����ڷۣ�ͨ���Dz���ʵ�ġ�������ʪ���漴���Ҳ���������ء����۵�����������ͷ��ά�ֶ�ã������ֹ�����һ���������յ��£������������������⣬�����ֲ����Ŵ���Ҳ�кܴ��ϵ��
�������������۹ܵ�����
����������ͷ�ϵĻ��ۣ��������ͷ�������������͵ģ�������ˮ��ø�Ĵٽ����ú㿪ʼ�ȷ����γɻ��۹ܡ����ڻ�������������ʼ�Ӳ����Χ���ڱ����ܣ�ֻ�����ȷ��ĵط�������չ��أ����Ի��۵�ԭ��������ڱڣ������͵�����£�һ������һ���ȷ���ͻ�����γ�һ��ϸ���Ĺ��ӣ���Ϊ���۹ܡ���Ȼ��Щֲ��Ļ��۾����ȷ��ף�������ơ���«��ֲ��Ļ��ۣ�����ͬʱ�����������۹ܣ���ֻ�����е�һ���ܼ���������ȥ���������;ֹͣ������
�������۹��ж������������ԣ���������ֻ����ǰ��3—5 μm�����γɺ��ܼ����������죬�ȴ�Խ��ͷ��Ȼ���������ӷ���ͬʱ��������ϸ�����ں���ȫ��ע�뻨�۹��ڣ��۹ܶ��˼��У��������ϸ���͵Ļ�������Ӫ���˺�2������ȫ�����뻨�۹��У�����ϸ���͵Ļ�������Ӫ���˺���ֳϸ�����뻨�۹ܺ���ֳϸ�����ڻ��۹��ڷ��ѣ��γ�2�����ӡ�
�������۹�ͨ�����������ӷ�������;�����ɷ�Ϊ���ֲ�ͬ�������һЩֲ��Ļ����м�ɿ��ĵĻ����������۹�������ʱ���Ż������������죬�����ӷ�����һ������ǻ���������������Ϊ�����������֯��transmitting tissue����һ�㱡��ϸ�������������۹�����ʱ�辭��ø�����ã���������֯����֯ϸ�����в�������ܽ⣬���۹ܾ���ϸ��֮��ͨ�������۹��ڻ����е������������û��۱��������������Ӫ���⣬Ҳ�ӻ�����֯��ȡ���ϣ���Ϊ�����ͽ��ɹܱںϳ�����֮�á����۹ܵ����������ڼ�˲��֣��뻨�۹ܶ���ԽԶ�IJ���Խ��˥�ϡ�
���������ȷ��ͻ��۹ܵ������ٶ��ڲ�ֲͬ����������������ı仯���Dz���ȫһ�µģ�����Ӵ��۵��ܾ���ʱ��Ҳ���ϴ�ľ��ֲ��Ļ��۹���������������ƻ���ɴ��۵��ܾ���ʱ��Ϊ5�죬����ֲ�����賤��һ���һ��ࣻ��һ��ũ����Ļ����ȷ����������ٶȽϿ죬����ˮ��ֻ��1.5Сʱ��С��1Сʱ��������15—32Сʱ����Щ������Ҫ���Ŵ�����������������֮�⣬�������ص�Ӱ��Ҳ��ʹ�ٶ������ı䣬�绨�������ĺû�������ʱ���µĸߵͺͿ��������ʪ�ȵȡ�
�������۹ܵ����ӷ��Ժ���ֱ��������ף��������ң�ֱ�����飩�����߾��������������������ڣ��������������飩��������������ң�ͳ��Ϊ����ܾ���porogamy��ͼ4��54��A����Ҳ�л��۹ܾ���������ĺϵ�������ҵģ���Ϊ�ϵ��ܾ���chalazogamv��ͼ4��54��B����ǰ����һ��ֲ�����У��������ټ��������ܡ����ҵ��ܾ�������һ���͡����⣬Ҳ�д����鱻���ɲ���������ҵģ����в��ܾ���mesogamy����������ټ������Ϲϡ����ۻ��۹���������ȡ����һ��;�����������ȷ��������������ң���һ����IJ���ԭ��һ����Ϊ�ڴ���ijЩ��֯�����������������������֯��̥�����ӷ��ڱں���ϸ���ȴ���ij�ֻ�ѧ���ʣ����յ����۹ܵĶ���������
��������������ֲ���˫�ܾ����̼�������ѧ����
�������۹ܾ��������������ӷ���ֱ�����飬Ȼ����ף������������ҡ���������֯�ϱ�����������۹ܿ��������������ң��������ĽϺ����������۹�����ͨ����ʵ��������֯�����ܽ������ҡ�
�������۹ܽ������ҵ�;���ڲ�ֲͬ���Dz�һ���ģ���������ϸ����һ����ϵ���д��Ѻ���ϸ��֮��������ҵģ��������д���1����ϸ���У�Ȼ��������ҵģ����ޣ������ƻ�1����ϸ����Ϊ�������ҵ�ͨ·�ģ������ÿ������Ǵӽ������ϸ������ģ������ס����۹ܽ������Һܵ�ĩ�˼������ѣ������Ӽ�����������ע�����ҡ�����ԭ������Ϊ�����������ڵĵ����������£�����ϸ�����Ʋ�Ϊ�Ի��۹���������ֱ�ӵ����ã������۹�����ϸ����ϸ���ʽӴ�ʱ������ѹ����ͻȻ�ı䣬���¹ܵ�ĩ�����ѣ�Ҳ����Ϊ���۹ܹܱڵ��ܽ⣬�緬�ѡ����飬Ҳ��ԭ��֮һ��

 ��
��
�������۹��е����������ͷŵ������к��ŷ������Ӻ���ϸ���Լ����Ӻ�2���˵��ںϡ�2�����е�1�������ںϣ��γ��ܾ��ѣ���ƺ��ӣ�����������Ϊ�ߡ���1�����Ӻ�2�����ˣ�������ˣ��ںϣ��γɳ�������ˣ��Ժ���Ϊ���顣��ϸ���ͼ���ͬʱ��2�����ӷֱ�����ںϵĹ��̣��DZ���ֲ��������ֳ����������Ϊ˫�ܾ���doublefertilization����ͼ4��55����
�����������ҵ�2�������Ƿ��ڽṹ���������Եȷ�����ȫ��ͬ��ͨ�����������Ĺ۲��о�֤����2���Ӽ����пɲ���IJ�����ڵġ�Russell��1980—1983�������ڵ����������о�������plumbagozeylanica������ʱ���۲쵽2�������е�1����ͻ������Ӫ��ϸ���ĺ��ϣ���1����ͨ��������˿���һ�������������⣬������Ӫ�����ϵ��Ǹ����Ӻ��϶������������壬����1�������϶�����壬��ˣ�������Ϊ�������ӿ����Ƕ��εģ�dimorphic����
��������ϸ����ϵľ��ӣ��ڽ�����ϸ�����Ѻ˽ӽ�ʱ�����˵�Ⱦɫ���������Ѻ˵ĺ�Ĥ�ϣ�Ȼ����ѷ�ɢ��ͬʱ����1��С�ĺ��ʣ��������˺��Ѻ˵�Ⱦɫ���������һ���ۺ˵ĺ���Ҳ�ʹƺ˵ĺ����ں���һ�𣬽�����һ�ܾ����̡���1�����Ӻͼ��˵��ںϹ��������������ӵ��ں��ǻ������Ƶģ����ӳ�ʱҲ�ʾ����Ĵ�״���Ժ��ɿ��뼫�˱���Ӵ���2��Ⱦɫ�ʺ�2���ʺϲ�������������̡����Ӻ��ѵĽ�ϱȾ��Ӻͼ��˽�ϻ��������Ծ��Ӻʹ����˵ĺϲ���ɵý��硣
�����ܾ�ʱ������ϸ����ϸ�����Ƿ������ϸ���У��Ӳ�ͬ�о�����ó��Ľ����Dz�ȫ��ͬ�ġ�һ����Ϊ����ϸ����Ҳ�����ܾ�����һ�ֳ��෴�۵㣬���������������о���֤����һ������Ϊ��ȷ�����������ܾ�ʱ�������ṹ��ʾֻ�����Եĺ˻����ںϣ�������ϸ������������ƻ�����ϸ���С�
�����ܾ��������еķ���ϸ����������Ѷ��������࣬��Ϊ�ߺ����鷢��ʱ�����ϣ������ȫ����ʧ��
��������ֲ���˫�ܾ���ʹ2��������Ĵơ��������ں���һ�𣬳�Ϊ1��������ĺ��ӣ��ָ���ֲ��ԭ�е�Ⱦɫ����Ŀ����Σ�˫�ܾ��ڴ����ױ��Ŵ��ԣ���ǿ������������������Ӧ�Է����Ǿ��нϴ������ġ��������ںϾͰѸ���ĸ�����в�����Ŵ����������һ���γɾ�˫���Ŵ��Եĺ��ӡ��������Ӽ���ͬ�����γɵĺ�������п����γ�һЩ�µı��졣���ܾ��ļ��˷�չ�ɵ�������������ģ�ͬ�����и���ĸ�����Ŵ����ԣ���Ϊ����һ�����ڵ����ϣ�����Ϊ���̺ͷ�չ��һ�ص��ṩ�������������ԣ�˫�ܾ���ֲ�����������ֳ�������������������ʽ��
�������ģ��ܾ���ѡ������
������ͷ�Ի��������ȷ����Լ����ҶԾ���ϸ���Ľ��룬������ѡ��������Ҳ����˵��ֻ���ܺ���ͷ������������������Э���Ļ������������ȷ�����ϸ��Ҳֻ�ܺ���������������Ӧ�ľ����ں���һ�����ԣ�����ֲ����ܾ���������ѡ���Եģ����ֶԻ��ۺ;��ӵ�ѡ���ԣ���ֲ���ڳ��ڵ���Ȼѡ�������±��������ģ�Ҳ�DZ���ֲ����������е�һ����Ҫ������ˣ���Ȼ������ͷ�Ͽ������в�ֲͬ��Ͳ�ֲͬ������Ļ��ۣ����ǣ�ֻ���ʺ�����һ�ܾ����̵�ֲ�ﻨ�ۣ����ܲ���Ч����
�����ܾ����õ�ѡ������Ϊ�������ע�⣬���������ָ�����ܾ��������û��ѡ���ԣ��Ͳ����ܱ��������ܾ��ͽ����ܾ��ĺ�����Ҳ�����ܵõ������ܾ����洦��ʵ��֤����������ò�ֲͬ�꣬������ͬ����Ļ�ϻ��۽����ڷۣ�ֻ�����ʺ�����ͷ�����ҵĻ����о����ȷ��Ŀ��ܣ������˽����Լ����ϵĻ���������ˣ����û���ڷۡ��˹������ڷ�����߲������˷��Խ���ԶԵ�ӽ��IJ����ԣ��Լ���ߺ���Ի�������Ӧ���������ܵ��ձ����ӡ�ѡ���ܾ�������Ϊũҵ����ʵ���������ĺô���ϣ����Ϊѡ�ֺ����ַ��������춨�˻�����
�����ڱ���ֲ���У�˫�����ѺͶྫ���ѵ��������Σ�Ҳ�з��֣����Ӿ��ӽ�����ϸ���ı�����ϸ����ͬ�����ã�ʹ�ߵ�Ӫ���������Ӵ����Ŵ��Է����仯��
�������壩���ں���ֳ����������
��������������£�����ֲ���������ֳ�Ǿ�����ϸ���;��ӵ��ںϣ��Է������ߣ�������Щֲ��������������ںϣ�Ҳ��ֱ�ӷ������ߣ�������������ں���ֳ��apomixis�������ں���ֳ��������ϸ���������ܾ���ֱ�ӷ������ߣ����ѹ�Ӣ������̵ȣ���������ƹ´���ֳ��part-henogenesis������������ϸ��������ϸ���˵ȷ���ֳ��ϸ���������ߣ���С��β�����߲ݵȣ�����������Ϊ��������ֳ��apogamy����Ҳ�е��������Ļ��鱻ϸ��ֱ�ӷ������ߵģ����������Citrus������Ϊ��������ֳ��apospory����
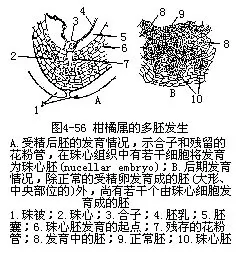
������һ������£�����ֲ�������ֻ����1�����ң�ÿ������Ҳֻ��1����ϸ���������ܾ���ֻ�ܷ�����1���ߡ����е�ֲ��������������2���������ߴ��ڣ���һ�����Ϊ��������polyembryony������������IJ�����������������������ֳ����������ֳ�Ľ����ͼ4��56����Ҳ��������1���ܾ��ѷ��ѳɼ����ߣ�����1�������з���������ҵ�Ե�ʡ�
��
������ ���Ӻ�ʵ
��������һ�����ӵ��γ�
��������������ʵ���γɺ�����
��������������ʵ�����ӶԴ�������Ӧ
��
������ ���Ӻ�ʵ
��
��������ֲ����ܾ�������ɺ�����㷢��Ϊ���ӣ�ͼ4��57�����ӷ�����Ϊ��ʵ����Щֲ������������ֺͻ�����Ľṹ��Ҳ������һ�����ɹ�ʵ��һ���ֵġ�
������������������ֲ�����е����١�����ֲ���е�����ֲ���Ϊ��������û�а������������鷢�������Ӻ�����¶�ģ�����ֲ��������ǰ����ӷ��ڣ���ϸ���ܾ����ӷ�����Ϊ��ʵ����������鷢�������ӣ���������Ҳ���ܵ���ʵ�İ������������ް�������������ֲ��������ֲ��ͱ���ֲ��������Ⱥ����Ҫ����֮һ������ֲ�������������ֳ�����ֵĸ��������⣬ͬʱҲ������ֲ����Զɹ��ɡ���Ȳ�����������Ч��ʩ������ʵ���ֳ����������⣬������������Ӫ����������ɢ�������á�
�����������ø���ֲ��Ĺ�ʵ��������Ϊʳ����ṩ��ҵ��ҽҩԭ�ϵȡ���ʳ�õĹ�ʵ���������༫�࣬�ճ�����������ȱ�ٵ���������ʳ�������ǵ��ס���ۡ��Ϲ������࣬���ֶ���ֲ������Ӻ�ʵ���֣����������һһ��������������ֲ��Ĺ�ʵ�������ǹ�ҵ�ϵ�ԭ�ϡ����Ӻ�ʵ�������صĵ��ۡ������ʡ���֬��������������ʳƷ��ҵ����֬��ҵ����ʳ�õĵ��ۡ�Ҭ�͡����͡����ͣ������ϵĿɿɡ����ȣ��Լ�����ҵ�õ������͡������͡�ͩ�͡������͡��������ȵȡ���ҽҩ�õĹ�ʵ����������Ҳ���٣�����顢�Ͷ���ʯ��ľ�ϡ�ʹ���ӵȡ���Щ�����²��н�ϸ��ơ���ֲ������������
�������¾����Ӻ�ʵ���γɡ��ṹ�����࣬�ֱ����������
��
��
һ�����ӵ��γ�
��
�������ӵĽṹ�����ߡ��������Ƥ�����֣��Ƿֱ����ܾ��ѣ����ӣ����ܾ��ļ��˺��鱻�������ɡ������ֲ������IJ��֣��������γɹ����У����������ö���ʧ��Ҳ��������������ļ���������ֱ�����ӳ��죬��Ϊ���ӵ������顣��Ȼ��ֲͬ�����ӵĴ�С����״���Լ��ڲ��ṹ���в��죬�����ǵķ������̣�ȴ�Ǵ�ͬС��ġ�

��
������һ���ߵķ���
������������������Ѿ����ܾ���ĺ��ӷ������ģ��������ߵĵ�һ��ϸ������ϸ���ܾ������һ����ά�ص�ϸ���ڣ���������״̬���ܾ�������ʱ�ڵij��̣���ֲ��������죬�н���Сʱ�ģ���ˮ�����ܾ���4—6Сʱ������һ�κ��ӷ��ѣ�С��Ϊ16—18Сʱ��Ҳ����2—3��ģ����ޣ��е����ӳ��������µģ���衢��ˮ�ɵȡ��Ժ��Ӿ���η��ѣ�����Ϊ���ӵ��ߡ�һ������£��߷����Ŀ�ʼ���ϳ�������ķ�����
����������һ���߶ȼ��Ի���ϸ�������ĵ�һ�η��ѣ�ͨ���Ǻ���ģ����������⣩����Ϊ����ϸ����һ��������ˣ���Ϊ��ϸ������һ��Զ��ģ���Ϊ����ϸ��������ϸ������Ϊ�ߵ�ǰ��������ϸ��ֻ��Ӫ���ԣ��������ԣ��Ժ��Ϊ�߱�����ϸ�����а�����˿��ͨ������ϸ���������ԣ����ɺ��ӵ����������������ġ�����û�г��ַֻ�ǰ�ĽΣ���ԭ�ߣ�proembryo������ԭ�߷�չΪ�ߵĹ��̣���˫��Ҷֲ��͵���Ҷֲ������в���ġ�
�����߱����ߵķ��������в�����һ�������ԵĽṹ����������ķ������߱�Ҳ�����ն���ʧ����ȥ���߱�����ʶֻ����Ϊ���Ű������������ڲ����ʵ�λ���������ڷ�����������Χ�����ϣ����������߱�ϸ���������ṹ�Լ���ֲͬ���߱������ޡ����̵ȷ���IJ��죬������Ϊ�߱�ϸ�������������һЩ���á��Ƚ�ͨ������ʶ�ǣ��߱������������⣬�����ܴ�������Χ����Ӫ��ת�˵����幩�������������Լ���Ϊ��Ҫ���ֺ͵��������������ʵĹ�ӦԴ��
����1��˫��Ҷֲ���ߵķ��� ˫��Ҷֲ���ߵķ�������������Ϊ��˵�������Ӿ��������ߺ����ȵغ�������Ϊ2��ϸ����������˵��ǻ�ϸ����Զ������Ƕ���ϸ������ϸ���Դ�����������ѣ��γ�һ����6—10��ϸ����ɵ��߱�����Щϸ��֮���а�����˿��ͨ�����������۲��߱�ϸ��������ͻ���������紫��ϸ����ϸ���ں���δ���ֻ������塣����ϸ����Ҫ���������ݷ��ѣ��ڶ��εķ��������һ�εĴ�ֱ������Ϊ4��ϸ�������ķ���ʱ�ڣ�Ȼ�����ϸ���ٺ������һ�Σ���Ϊ8��ϸ������״�壬���˷��壨octant��ʱ�ڡ��˷���ĸ�ϸ���Ƚ���һ��ƽ�ܷ��ѣ��پ�������������������ѣ���Ϊһ����֯�����ϸ���ʱ�ڶ���ԭ�߽Ρ��Ժ�����������֯�Ķ���������������Ͽ죬�γɶ���ͻ��Ѹ�ٷ�������Ϊ2Ƭ��Ҷ��������Ҷ��İ��ݲ����ֻ�����ѿ�����ͬʱ�����������·����߱�����һ��ϸ�������߸�ԭϸ����hypophysis��������������Ļ���ϸ��Ҳ���Ϸ���������һ��ֻ�Ϊ�߸����߸�����Ҷ��IJ��ּ�Ϊ���ᡣ��һ�ε����壬�������濴�����ٳ������Ρ����ã�����ϸ���ĺ�����ѣ�ʹ��Ҷ�������ӳ������������Ҷ���ڿռ��λ������Ҳ�����������Σ�ͼ4��58�������ˣ�һ�������������Ѿ��γɣ��߱�Ҳ���˻���ʧ��
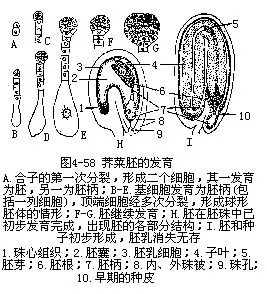 ����
����
����2������Ҷֲ���ߵķ��� ����Ҷֲ���ߵķ��������Ժ̱��Ƶ�С��Ϊ��˵����С���ߵķ�������˫��Ҷֲ���ߵķ�������й�ͬ������Ҳ�����𡣺��ӵĵ�һ�η�����б��ġ���Ϊ2��ϸ��������2��ϸ���ֱ���Խ���һ��б��ķ��ѣ���Ϊ4ϸ����ԭ�ߡ��Ժ�4��ϸ���ָ��Բ��ϵشӸ���������ѣ�������������������16—32ϸ��ʱ�ڣ��߳��ֹ���״���ϲ����Ϊ�����ǰ�����²�ϸ�����ֻ�Ϊ�߱�������������Χ��һ��ԭ��Ƥ��ϸ������Χ��
�������ã��ڰ�״�����һ�����һ��С�ΰ��̣����ڰ��̴��γ���������������㣬�������ϵ�һ�������巢չΪ��Ƭ����Ҷ����������һ���������Ͽ죬���Ժܿ�ͻ����������֮�ϡ�������ֻ��ã���������ѿ�ʵ�ԭʼ�壬��Ϊһ���۵���֯������������͵�һƬ��Ҷԭ�������档���ͬʱ�����������Ҷ��Ե���һ�࣬�γ�һ���µ�ͻ�𣬲���������Ϊ����Ҷ��������Ҷ��������ϸ���ľӼ�������������Ҷ�ϲ��쳤�ܿ죬���ó�Ϊ��Ƭ�������ߵ�һ�ࡣ
������ѿ�ʿ�ʼ�ֻ����ֵ�ʱ����������·������߸��ʺ��߸���ԭʼ�壬�����߸����߸���ϸ���������ٶȲ�ͬ���������߸���Χ�γ�һ�������ԵĿ�ǻ�������ߵij���ǻҲ���ϵ�����
�������ˣ�С��������ѻ����Ϸ����γɡ��ڽṹ�ϣ�������һ�Ŷ�Ƭ����Ҷ����λ���ߵ��ڲ࣬���������������������������Լ���һƬ��Ҷԭ���ϳ���ѿ����������ѿ�ʰ������������ѿ��һ�����߸��������߸��ʰ����������Ƭ��Ե�һ�棬���Լ�������Ҷ��ͻ��ͼ4-59�����еḻ́���ֲ���������ߣ�����������Ҷ��
 ��
��
��������������ķ���
���������DZ���ֲ�������������ϵIJ��֣���2�������ܾ��������ɣ������������ںϣ�triplefusion���IJ�������ܾ��������ߣ���������ϸ�����������顣
��������ķ�����һ���к��ͣ�nucleartype��non-cellulartype����ϸ���ͣ�cellulartype��������Ŀ�ͣ�helobialtype�����ַ�ʽ���Ժ��ͷ�ʽ��Ϊ�ձ飬������Ŀ�ͱȽ��ټ���ֻ����������Ŀ��Helob-iales��ֲ������鷢���С�
������������ķ������ܾ����˵ĵ�һ�η��ѣ��Լ����һ��ʱ�ڵĺ˷��ѣ�������ϸ���ڵ��γɣ�����ϸ���˱�������״̬���ֲ���ͬһϸ�����У���һʱ�ڳ�Ϊ����˵��γ��ڣ�freenuclearformationstage��������У����Ŀ����ֲ��������죬��Ŀɴ�����������ǧ�����Ź��ɵ�ϸ��ʱ�ڣ�����ҡ�ƻ���ȡ��ٵĽ�8��16���ˣ�����ֻ��4���ˣ��翧�ȡ����ź��������ӣ��˺�ԭ��������������Һ�ݵij��֣������������ҵ����ܣ������ҵ���˺ͺϵ�˽�Ϊ�ܼ����������ҵIJ���ֲ���һ���㡣�˵ķ�������˿���ѷ�ʽ����Ϊ�࣬Ҳ������������˿���ѣ��ر����ںϵ�˷ֲ��ĺˡ�
��������˷��ѽ��е�һ���Σ�����ϸ��ʱ�ڹ��ɣ���ʱ�������֮���γ�ϸ���ڣ�����ϸ���ʵķָ������γ�����ϸ����������֯��Ϊ���顣�������鷢����ֲ��������е���ȫ������˶�תΪ����ϸ����Ҳ�н����ҵ���Χ�γ�һ����ϸ�����������Ա��������״̬������ϸ��ֻ���������ҵ�����γɣ������������Dz��γ�ϸ���ġ�����Ҷֲ��Ͷ���˫��Ҷֲ��������һ���ͣ�ͼ4��60����
 ����
����
����ϸ��������ķ�����ͬ��ǰ�ߵĵط������ں˵�һ�η��Ѻ��漴����ϸ���ʵķ��Ѻ�ϸ���ڵ��γɣ��Ժ���еķ���ȫ��ϸ�����ѣ�����������ʼ������ϸ������ʽ�������������ʱ�ڣ���������Ϊ��ϸ���ṹ��������ϰ껨��ֲ��������һ���ͣ�ͼ4��61����
��������Ŀ������ķ������Ǻ��ͺ�ϸ���͵��м����͡��ܾ����˵�һ�η���ʱ�����ұ���Ϊ2�ң�������Һͺϵ��ҡ�����ұȽϴ���һ���ֵĺ˽��ж�η��ѣ�������״̬���ϵ��Һ˵ķ��Ѵ������٣���һֱ��������״̬���Ժ�����ҵ�������γ�ϸ���ṹ���������ķ�����������һ���鷢�����͵�ֲ�����������Ŀ���࣬��̹���к���ȹá���β������Eremurus sp.���ȣ�ͼ4��62����
������Ϊ�����������ںϵIJ��������2�����˺�1�����Ӻˣ�������������Ⱦɫ�壨��ĸ���ṩ2���������ṩ1���������ԣ���ͬ�������Ÿ�����ĸ��ֲ����Ŵ��ԡ��������������巢�������е����ϣ�Ϊ�����������ã���ˣ����߷������Ӵ������Ը�����������ǿ����Ӧ��Ҳ���㡣


������������������ϣ��еľ�����������ȷ�ʱ��Ϊ�������õģ��������������飬��Ϊ���������ӡ���ǰ���ᵽ���ḻ́���ֲ�����ӡ��������ӵȡ�������һЩֲ������ߵ��γɣ������漴�������գ����浽��Ҷ��������ӳ���ʱ����������ڣ���Щ�����������ӣ��綹�ࡢ��������ӡ�
����һ��ֲ�����ӣ����ߺ����鷢�������У�Ҫ����������Χ������֯�����ϣ���������һ����ƻ�����ʧ��������ֲ�����������ʼ�մ��ڣ����������з����������������һ��Ӫ��������֯����Ϊ�����飨prosembryum�������������������ã�����Դ�����鲻ͬ��������������ӣ�������������ṹ�ģ���������ʯ��������˵ȣ�Ҳ������������ṹ�ģ�����������ȡ�
��������ֲ���е����ơ���̦�ݿơ���Ƶ�ֲ������ڷ��������м�����Ҳ�����ܾ����ã����ܾ����˲����˻���ʧ����������Ϊ���飬���������ڲ���������ṹ��
�������������ӵ��γ�
�������ӵ������һ��Ϊ��Ƥ����������Ƥ����������鱻�����ߺ����鷢����ͬʱһ�������ɵġ��鱻��1��ģ�Ҳ��2��ģ�ǰ�߷����ɵ���Ƥֻ��1�㣬�����տ������ң����߷����ɵ���Ƥͨ��������2�㣬������Ƥ������Ƥ��������������ȡ���������ֲ���У�һ�����鱻����֯��Ӫ���������գ�����ֻ��һ���ֵ��鱻��Ϊ��Ƥ���е����ӵ���Ƥ����2���鱻�е����鱻�������ɣ�����϶���Ҳ�е���2���鱻�е����鱻���������ģ���ˮ����С��ȡ�����ֲ�����ӵ���Ƥ�����Ǹ���ģ���Ҳ���������������ʵģ���ʯ�����ӵ���Ƥ�������ƤΪ��֭��ϸ��������ɣ������ӵĿ�ʳ���֡�������Ƥ������ֲ����������dz����ģ����ӵ�����Ƥ���Ƿʺ����ʵġ�
������Ƥ�Ľṹ������ֲ�����ϴ�һ����������鱻����Ŀ��ͬʱҲ��������Ƥ�ڷ����еı仯��Ϊ���˽���Ƥ�ṹ�Ķ������������Բ϶����Ӻ�С��������Ƥ�ķ������Ϊ��������˵����

 ��
�������϶��������γɹ����У���������鱻Ϊ���������ģ������������ڣ�������Ƥ�������鱻����֯��չ���ġ����鱻��������Ƥʱ���鱻�ֻ�Ϊ3����֯�����ϸ����1�㳤��״���ϸ����ϸ���ij������ܵ�ƽ�����У�����դ״��֯���ڶ���ϸ���ֻ�Ϊ���κ��ϸ������Щϸ������״����������̿���“��”���Σ��ں�ϸ��ǻ���ԣ��˴˽������У��м�ǿ�ı������úͻ�е�������������Ƕ�㱡��ϸ���������鱻δ���ֻ���ϸ���㣬�����ڳɳ�ʱ���ⲿ��ϸ������ѹ�⡣���ڵ���Ƥϸ���ں��е��ۣ���Ӫ������ij����������������۵IJ϶���Ƥ������ʳ���Ϻ��תΪ���͵���֯��ͼ4-63����
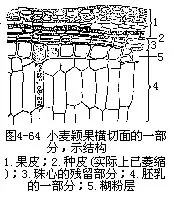
����С�����ӷ���ʱ��2���鱻Ҳͬ������һϵ�б仯����ʱ��ÿ���鱻������2��ϸ�������ӽ��е�һ�η���ʱ�����鱻��ʼ�����˻�����ϸ����ԭ��������ʧ���Ժ���ʧȥԭ��ϸ����״��������ʧ�����鱻��ʱ�б���ԭ����״�����������������������ʱ�ڣ����鱻�����ϸ����ʼ��ʧ���ڲ�ϸ�����ֶ��ڵĴ��ڣ������ӳ������ʱ�����������˱������ã��Ժ���Ϊ���ӱ�������֯�㣬��Ҫ������Ƥ����������ͼ4-64����
��
������ʵ���γɺ�����
��
�����ܾ������Ժ��ĸ��������������ı仯�����ࣨ�����������⣩������һ���ή���䣬����ʹ������ͷ������Ҳ����л�����ӷ������ӷ�����������֮�����IJ��֣�Ѹ�������������ɹ�ʵ��һ����ԣ���ʵ���γ����ܾ���������������ϵ����ֻ�����ܾ�������γɹ�ʵ�������е�ֲ������Ȼ״������Ϊ���Ƶ������£��䲻�����ܾ����ӷ�Ҳ�ܷ���Ϊ��ʵ�������Ĺ�ʵ���治�����ӡ�
������ʵ�����ʺͽṹ�Ƕ��ֶ����ģ����뻨�Ľṹ���ر�����Ƥ�Ľṹ���Լ��ܾ�����Ƥ�����������ֵķ���������кܴ��ϵ�������ʵ���γɡ��ṹ�Լ����ͣ��ֱ����������
������һ����ʵ���γ�
����1����ʵ���γɺͽṹ��ʵ�е������ӷ��������ɵģ�Ҳ�����ɻ������������绨�С����ࡢ�������һ�������ɡ�
�����ӷ�ԭ���ɱ���ϸ�������ɣ��ڷ����ɹ�ʵʱ������һ���ֻ�Ϊ���ֲ�ͬ����֯���ֻ���������ֲ��������죬���ǹ�ʵ��������ݡ�
������ɹ�ʵ����֯����Ϊ��Ƥ��pericarp����ͨ���ɷ�Ϊ����ṹ������������Ƥ��exocarp�����в����й�Ƥ��mesocarp�����ڲ����ڹ�Ƥ��endocarp���������Ƥ�ĺ�Ȳ���һ�µģ��ӹ�ʵ������졣��Щ��ʵ������Ƥ�ֽ�Ƚ����ԣ�������еĺ˹��ࣻҲ�зֽ粻����ȷ�����������ϣ�������ġ���Ƥ�ķ�����һ��ʮ�ָ��ӵĹ��̣��������ܵ����غ��ӷ��ڵ��ڡ��С������֯��Ӧ������������������Ƥ����֯�㣬���ڷ��������г��ַֻ���ʹ�����ǵ���Դ���Ե����ѡ�
�����ϸ��˵����Ƥ��ָ������ӷ��ڣ������ʵ����ɲ��֣�����Ƥ�⣬�а������������ṹ��֯�ģ��绨�еȣ����Ƥ�ĺ���Ҳ�������ӷ��ڵĸ����ṹ����֯���֡�
����2�����Խ�ʵ�����ӹ�ʵ��ʵ���γɣ�һ�����ܾ����������й�ϵ����Ҳ�в����ܾ����ӷ��ͷ����ɹ�ʵ�ģ��������γɹ�ʵ�Ĺ��̳Ƶ��Խ�ʵ��parthenocarpy�������Խ�ʵ�Ĺ�ʵ�ﲻ�����ӣ����Գ������ʵΪ���ӹ�ʵ��
�������Խ�ʵ���Է��γɵģ���Ϊ�Է����Խ�ʵ��autonomous parthenocarpy����ͻ�����������㽶���㽶�Ļ�������״�����������ϲ����ۻ����²��Ǵƻ����ƻ��ɲ������ۡ��ܾ����γɹ�ʵ����������Ȼ�������ܽ��е��Խ�ʵ�������ѵ�ijЩƷ�֡����١���������ȣ���Щ����ֲ��Ĺ�ʵ�в������ӣ�Ʒ�������������ϵ���������Ʒ�֣���һ�������ͨ��ij���յ������������Խ�ʵ�����յ����Խ�ʵ��induced parthenocarpy�����������������Ļ��۴̼����ѵ���ͷ��������ɽ���Ļ��۴̼����ѵ���ͷ�����ܵõ����ӹ�ʵ���������ø��������̼���Ϳ�����������ͷ�ϣ�Ҳ�ܵõ����ӹ�ʵ������������2��4-D������������������أ��ڹ���ͷ������յ����Խ�ʵ��ȡ������Ч����
������������ʵ������
������ʵ�����Ϳ��ԴӲ�ͬ���������֡���ʵ�Ĺ�Ƥ�������ӷ��ڷ������ɵģ���Ϊ�����true fruit��������ֲ��Ĺ�ʵ����һ��������ӷ��⣬�����������ֲ����ʵ��ɵģ��绨�����������������ᣬ�����ʵ��Ϊ�ٹ���spurious fruit��false fruit������ƻ�������ࡢ����ȡ�
�������⣬һ�仨�����ֻ��һö����Ժ�ֻ�γ�һ����ʵ�ģ���Ϊ������simple fruit�������һ�仨����������������Ժ�ÿһ�����γ�һ��С���������ͬһ����֮�ϣ���Ϊ�ۺϹ���aggregate fruit������������ݮ�������ӵȣ�ͼ4-65���������ʵ������������������������Ҳ�����ʵ����ɲ��֣���ͳ�Ϊ�ۻ�����collective fruit����ƻ������Ҳ�Ƹ�����multiplefruit������ɣ�����桢�����ȣ�ͼ4��66����
�����������Ƥ�����������֣��зʺ����ʵģ��������fleshy fruit����Ҳ�й�ʵ�����Ƥ������֭�ģ��Ƹɹ���dry fruit��������ɹ��ָ������������͡�

 ��
��
����1����������ǹ�Ƥ���ʻ��������ʺ��֭���ڳ�������г�����һϵ�������仯���磺�����ɵ���ת���ɿ������ǣ��л�������������ࣻ����Ҳ�������Ϊ����״̬���Ӷ������˹�ʵ����ζ��������ζ��ɬζ�������е�Ҷ�����ƻ���ϸ��Һ���ֻ����أ�ʹ��ʵ����ɫ����ת�䣻����ϸ���в���ijЩ�ӷ���֬�����ʣ�ʹ��ʵ���㣻����ϸ���İ�������ڹ���ø�����ö��ܽ⣬ʹ������������Ϊɫ���㡢ζ���汸�Ŀ�ʳ�ò��֡�����ֿɰ���Ƥ��Դ�����ʲ�ͬ����Ϊ���¼��ࡣ
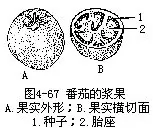
������1��������berry���������������Ϊϰ����һ�࣬��һ������Ƥ�γɵĹ�ʵ����Ƥ�����漸��ϸ���⣬һ�����ۣ����ʶ���֭���ں��������ӣ������ѡ����ѡ����ȡ����ѹ�ʵ������ʳ�ò��֣���Ҫ���ɷ����̥����չ���ɣ�ͼ4��67����
�� ��
��
������«��ֲ��Ĺ�ʵ������ֹ��࣬�ǽ�������һ�֣�һ���Ϊ����pepo������ʵ�����ʲ������ӷ��ͻ��й�ͬ�������ɵģ��������ڼٹ����Ϲϡ����ϵ�ʳ�ò��֣���Ҫ�����ǵĹ�Ƥ�������ϵ�ʳ�ò�����ԭ����̥����
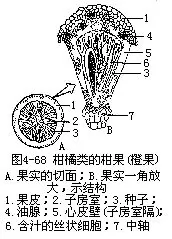
����������Ĺ�ʵҲ��һ�ֽ������Ƴȹ���̹���hesperidium�������ɶ���Ƥ������̥�����ӷ��������ɡ��������Ƥ�����ʣ��кܶ����ҷֲ����й�Ƥ�������ʣ���ά�����ֲ���䣬�����Ƥ�ڵ�“����”������Щά�������ڹ�ƤĤ�ʣ���Ϊ�����ң����ڳ�����֭�ij���˿״ϸ������ԭ���ӷ��ڱڵ�ë�������ɣ��������ʵ��ʳ�ò��֣��糣���ĸ��١��֡����ʵȣ�ͼ4��68����
������2���˹���drupe��ͨ���ɵ����չ���ɣ��ں�һö���ӣ������Ƥ���ʲ�һ�����Ƥ���������ӷ���Ƥ�ͱ�Ƥ�¼���ϸ����ɣ��й�Ƥ�Ƿ��������ʳ�ò��֣��ڹ�Ƥ��ϸ����ľ�ʻ���Ϊ��Ӳ�ĺˣ������������棬���ֹ�ʵ��Ϊ�˹���ͼ4��69�������ҡ�÷����ӵȵĹ�ʵ��Ҳ�г���ĺ˹��й�Ƥ������֭�ģ���Ҭ�ӡ�Ҭ�ӵ��й�Ƥ����ά״���׳�Ҭ�أ��ڹ�Ƥ��ΪҬ�ǡ�

 ��
��
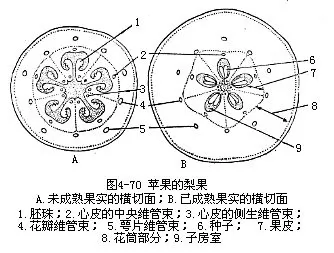
������3�������pome�������ʵ��Ϊ���ӷ���λ����ֲ�����С���ʵ�ɻ�Ͳ����Ƥ�������Ϻ�ͬ�γɣ�������һ��ٹ�������ܺ�����ʲ�����ԭ���Ļ�Ͳ�����ʲ������ڲ��ǹ�Ƥ���֡����Ƥ�ͻ�Ͳ���Լ����Ƥ���й�Ƥ֮�䣬�������Խ��ɷ֡��ڹ�Ƥ��ľ�ʻ��ĺ��ϸ������ɣ����ԱȽ�������ԡ��桢ƻ�����������ʵ�ĵ��ʹ�����ͼ4��70����
����2���ɹ���ʵ�����Ժ�Ƥ����еĹ�Ƥ�����п��ѣ�Ϊ�ѹ���Ҳ�м�ʹ��ʵ���죬��Ƥ�Ապϲ����ѵģ�Ϊ�չ���������Ƥ�ṹ�IJ�ͬ���ֿ�����Ϊ���¼������͡�
������1���ѹ��ࣨdehiscentfruit����ʵ������Ƥ�����ѿ����ɷ�Ϊ���¼������ͣ�
�����ټԹ���legume���Թ��ǵ���Ƥ�������ɵĹ�ʵ�������Ƥ�ر���������濪�ѣ�����㶹���϶��ȣ��е���Թ���ʽ�����������ѣ����仨�����ϻ������Եȣ�Ҳ�еļԹ��ʷֽ�״�������Ҳ�����ѣ����ǽڽ����䣬ÿ�ں�����һ��������Թ�����Ϊ�ڼԣ�����������߲ݡ�ɽ��ȵȣ��еļԹ�����״�����д�ë������ޣ�Ĺ�ʵ����Բ���ηֽڣ�������״���绱�Ĺ�ʵ��ͼ4��71����

��
�������uȋ����follicle���uȋ�����ɵ���Ƥ��������Ƥ�������ɵĹ�ʵ�������ֻ��һ�濪�ѡ�������Ƥ���쿪�ѵģ�����ͩ��ĵ������ҩ���˽�����ȵĹ�ʵ��Ҳ���ر��쿪�ѵģ���ľ�����������ȣ�ͼ4��72����
 ����
����
������������capsule���������ɺ�����Ƥ�ĸ���������ɵĹ�ʵ���ӷ���һ�ҵģ�Ҳ�ж��ҵģ�ÿ�Һ����Ӷ����������ʵ��Ϊ�ձ飬����ʱ�����ַ�ʽ���ѣ���1�����ѣ�longitudinaldehiscence�����ѷ�����Ƥ���᷽��ֿ����ֿɷ�Ϊ���Ҽ俪�ѣ�septicidaldehiscence����������Ƥ������Ӵ��ѿ�������ʱ�ɶ�������Ƥ��ϵ��ӷ���Ĥͬʱ�ֿ�������ˮ�ɡ������塢�����ȣ��ұ����ѣ�loculicidaldehiscence��������Ƥ���촦���ѣ����β�����ޡ������ݡ��ϻ��ض��ȣ����Ὺ�ѣ�septifragaldehiscence��������Ƥ�����Ҽ���ұ����ѣ����ӷ���Ĥ����������������ǣţ����ܡ������ȡ���2�����ѣ�porousdehiscence������ʵ�������Ƥ�������룬�����ӷ������Ϸ��ѳ�С�ף������ɿ�ɢ��������ڡ�����ݡ��۹��ȡ���3�����ѣ�circumscissiledehi-scence����������Ƥһ�ҵĸ�������ɣ���Ƥ������ع�ʵ���ϲ����в������ѣ���ʵ�ɸ�״���ѣ���ӣ�ݡ������ȡ���ǰ�ȣ�ͼ4��73����Ҳ�Ƹǹ���pyxis����
 ����
����
�����ܽǹ��ǹ�����2��Ƥ��ɵĴ�������ɵĹ�ʵ���ӷ�1�ң���������Ƥ��Ե������������������Ĥ�����ӷ��ָ���2�ң���һ��Ĥ����Ϊ�ٸ�Ĥ����ʵ�����Ƥ�ɻ���������2�����ѿ�����2Ƭ���䣬ֻ���ٸ�Ĥ�����Ӹ��ڼٸ�Ĥ�ϡ�ʮ�ֻ���ֲ���������ʵ���ǹ���ϸ���ģ����������ö����Ϊ���ǹ���silique������̦ܿ���ܲ��������ȣ�����һЩ���εģ�����֮�ȼ���ȣ���Ϊ�̽ǹ���silicle���������ˡ������˵ȣ�ͼ4��74����
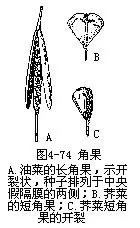
�� ��
��
������2���չ��ࣨindehiscentfruit����ʵ�����Ƥ�Բ����ѣ��ɷ�Ϊ���¼������ͣ�
�������ݹ���achene����1—3��Ƥ���ɵ�С�ͱչ�����Ƥ��Ӳ�����ں�1ö���ӣ�����ʱ��Ƥ����Ƥ����һ�����������ڷ��롣���ͷ�̣�1��Ƥ���ɣ������տ����ѹ�Ӣ��2��Ƥ���ɣ�������3��Ƥ���ɣ���ͼ4��75��A���ȡ�
������ӱ����caryopsis��ӱ���Ĺ�Ƥ�������ʣ�ֻ��һ�����ӣ���Ƥ����Ƥ�������ϲ����롣��ʵС��һ��������Ϊ���ӣ���ˮ����С�����Ⱥ̱���ֲ������й�ʵ���ͣ�ͼ4��75��B����
�����۳����samara������Ĺ�ʵ�������ݹ����ʣ�����Ƥ��չ�ɳ�״�����������Ʈ�ɣ����ܡ��ʡ�������ֲ��Ĺ�ʵ��ͼ4��75��C��D����
�����ܼ����nut����������Ƥ��Ӳľ�ʣ���һ�����ӵĹ�ʵ�������ʵ���ԭ������ܰ�����Ϊ�Ƕ������ݡ�黺����ȹ�ʵ��ͨ��һ�������н���һ����ʵ���죬Ҳ��ͬʱ�ж�������ʵ����ģ���������������������̵Ŀǣ������������ڹ�ͬ�Ŀ��ڣ������ɻ����ܰ��������ɣ�ͼ4��75��E����
������˫������cremocarp��˫��������2��Ƥ���ӷ��������ɵĹ�ʵ��ɡ�ο�ֲ��Ĺ�ʵ��������һ���͡��������Ƥ��������꣬��������������������϶ˣ�����������Ƥ�У��Ժ����롣��Ƥ��������ѣ�����ܲ���С����Ĺ�ʵ��
�����ް�����utricle�����“�ҹ�”�����ɺ�����Ƥ�γɵ�һ���ʵ����1ö���ӣ�����ʱ����������ѡ���Ƥ�������ɵذ�Χ���ӣ����������ӷ��룬���������ط��ȵĹ�ʵ��
��
������ʵ�����ӶԴ�������Ӧ
��
��������ֲ�����Է�ֳ�����нṹ——���ӣ��ǰ��ڹ�ʵ���ܹ�ʵ�����ģ�ͬʱ����ʵ�ĽṹҲ���������ӵ�ɢ������ʵ������ɢ�����أ�������ֲ���������Χ���뷱�������������ģ�ҲΪ�ḻֲ�����Ӧ���ṩ������
������ʵ�����ӵ�ɢ������Ҫ����������ˮ��������������Я�����Լ�ͨ����ʵ�����������Ļ�е��������ʵ�����Ӷ��ڸ���ɢ����������Ӧ��ʽ�Dz�һ���ģ��ֱַ��������¡�
������һ���Է���ɢ������Ӧ
��������ֲ��Ĺ�ʵ�������ǽ�������ɢ���ģ�����һ��ϸС���ᣬ�������ڿ�����Ϊ�������͵�Զ����������ֲ�������С���ᣬ����紵�͵�����������ķ�Χ�ڷֲ�������ǹ�ʵ�����ӵı��泣������ë�����ᣬ�����������ڳ��ܷ������������졣���ޡ������������涼��ϸ������ë�����������������ѹ�Ӣ��ʵ�ϳ��н���ɡ״�Ĺ�ë����ͷ�̹�ʵ�ϴ��������״��ͷ���ʡ��ܵȵĹ�ʵ�Լ��ɡ���ɼ�����ӵ�һ���ֹ�Ƥ����Ƥ��չ�ɳ�״�������Ὤ����Physalis���Ĺ�ʵ�б�Ĥ״�����ң���Щ�������ڷ������͵����нṹ��ͼ 4��76�����ڲ�ԭ�ͻ�Į�ϵķ���ݣ�tumble weed�������ӳ���ʱ�����ε�ֲ���ڸ��������룬��紵�����ֲ�����Զ�ij�����������ë������Salsola����˿ʯ������Gypsophila���ȡ�
 ����
����
������������ˮ��ɢ������Ӧ
����ˮ���������������ֲ���ʵ������������ˮ�����͡����Ĺ�ʵ���׳�����ʵ�Բ�Σ���֯���ɣ����ᣬƮ��ˮ�棬��ˮ����������ͬʱ������Զ�����أ�ͼ4-77����½��ֲ���е�Ҭ�ӣ����Ĺ�ʵҲ�ǿ�ˮ��ɢ���ġ�Ҭ�����й�Ƥ���ɣ�������ά����Ӧ��ˮ��Ʈ�����ڹ�Ƥ�ּ���ɷ�ֹˮ����ʴ����ʵ�ں�����Ҭ֭������ʹ�߷��������ʹҬ��������ˮ�Ļ����������ȷ����ȴ������ش���Ҭ�ֲַ������ʵ��ɢ������һ����ϵ�ġ�

 ��
��
�����������Զ��������ɢ������Ӧ
����һ����ֲ��Ĺ�ʵ�������ǿ�����������Я��ɢ�����ģ������ʵ�����ӵ��������д�ë����������ճҺ���ڣ��ܹ��ڻ�ճ���ڶ����ë���𣬻����ǵ��¿��ϣ����Ŷ�������ǵĻ�����а�����ɢ������Զ�ĵط��������¡�����ݡ��Զ������������꣨ͼ4��78���͵��Ρ����вݵȡ�
������ʵ�еļ��������ijЩ�����ʳ�ϣ��ر��������������ʵ���˿�ȥ����ص��£���һ���ֱ��Ե��⣬����ľ���ԭ�������ȷ����������϶�һЩС��ֲ������ӣ�Ҳ�����ƵĴ�����ʽ��
�������ڹ�ʵ�е�����࣬�����������ϲ����ʳ�ϣ���Щ��ʵ����ʳ��Ƥ���ֱ��������գ����������ӣ����ڼ�����Ƥ�ı��������������������ķ���ų���ɢ���������������ʺϣ������ȷ���ͬ��������ֲ��Ĺ�ʵҲ�������ճ������еĸ���ʳƷ����ȡʳʱ�����������洦���������ӽ��ȡ���˹�Ϊɢ���Ļ��ᡣ
�������ģ�����ʵ�����Ļ�е����ʹ����ɢ������Ӧ�ṹ
������Щֲ��Ĺ�ʵ�ڼ��翪��ʱ��������е��������������ʹ����ɢ����ȥ���ɹ��е��ѹ��࣬��Ƥ������Ϊ�����Ӳ�Ľṹ�����ڹ�Ƥ������ϸ����������ʽ��һ�����Ź�Ƥ��ˮ���ı仯������������ʱ����Ť������˰����ӵ�������ɢԶ���������Ĵ��϶������ɻ��ȹ�ʵ�д��������Դ��Ͳ˵Ⱦ���ֲ��Ĺ�ʵ���������뼰ʱ�ջ�Ȼ����������п��ѣ�������ɢ������䣬������ʧ����ϵĹ�ʵ����ʱ���ڶ����γ�һ���ѿף�����ʵ����ʱ���ɽ������絽Զ����ͼ4��79����
��

���߽� ֲ�������ʷ
��
��������ֲ���ھ���һ��ʱ�ڵ�Ӫ�������Ժ������ֳ�Σ���ʱ��ֲ�����һ����λ�γ���ֳ�ṹ��������ֳϸ�����з�ֳ������������ֳ�����γ������壬�����Ѻ;��ӣ��ںϺ��γɺ��ӣ�Ȼ�������µ�һ��ֲ���塣��������ֲ����һ�����������ķ����ͷ�ֳ�Σ�ǰ����̣��й��ɵ�ѭ����ȫ�����̣���Ϊ����ʷ��lifehistory�����������ڣ�life ycle����
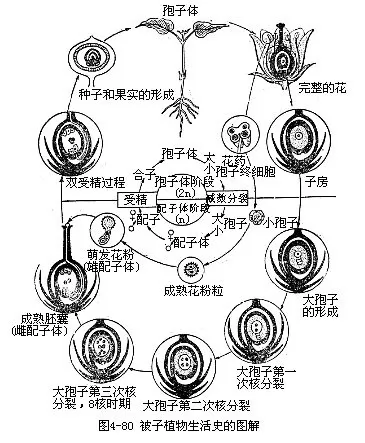
��������ֲ�������ʷ��һ����Դ�һ�����ӿ�ʼ���������γ��Ժ���һ�����ݵ������ڣ��ڻ���ʺϵ����ں���绷������ʱ�����ȷ�Ϊ���磬�����ɾ߸�������Ҷ��ֲ���塣����һ��ʱ�ڵ����������Ժ�һ���ֶ�ѿ��Ҹѿ���ٷ���Ϊ֦��������ת��Ϊ��ѿ���γɻ��䣬������Ļ�ҩ�����ɻ������������ӷ����������γ����ҡ��������������ָ��Էֱ�������Ծ��Ӻʹ��Ե���ϸ�����������ۡ��ܾ���1�����Ӻ���ϸ���ںϣ���Ϊ���ӣ��Ժ��������ӵ��ߣ���1�����Ӻ�2�����˽�ϣ�����Ϊ�����е����顣����ӷ�����Ϊ��ʵ�����鷢��Ϊ���ӡ���������������������һ���ij��塣��ˣ�һ���“�����ӵ�����”��һȫ�����̣���Ϊ����ֲ�������ʷ���������ڡ�����ֲ������ʷ��ͻ���ص�����˫�ܾ���һ���̣�������ֲ����û�еġ�
��������ֲ�������ʷ���������������Σ�һ���Ƕ�����ֲ��Σ�2n����һ���֮Ϊ������Σ�����Ǿ߸�������Ҷ��Ӫ����ֲ�ꡣ��һ���Ǵ��ܾ��ѷ�����ʼ��һֱ����������Ĵ�����ֱ��γ�����ĸϸ����������ĸϸ�����ͻ���ĸϸ����С����ĸϸ�������м�������ǰΪֹ������������ֲ������������У�ռ�˾��ֵ�ʱ�䡣��һ��ֲ����ĸ�����ϸ��Ⱦɫ�������Ƕ����ġ��������Ҳ��ֲ��������ԽΣ�����Ҳ��Ϊ������������һ���ǵ�����ֲ��Σ�n����һ��ɳ�Ϊ������Σ�������������������ɴ�����ĸϸ�������������Ѻ��γɵĵ��������ң������ӣ�����С����ĸϸ�������������Ѻ��γɵĵ����ڻ���ϸ����С���ӣ���ʼ��һֱ�����ҷ����ɺ���ϸ���ij������ң��ͻ��۳�Ϊ��2������3����ϸ���ij��컨���������ȷ��γ����������ӵĻ��۹ܣ���˫�ܾ�����Ϊֹ������ֲ�����һ��ռ������ʷ�еļ���ʱ�ڣ����Ҳ������������ֲ��������档�ɾ����ں����ɺ��ӣ�ʹȾɫ���ָֻ����������������������½��뵽������Σ������һ���������ڡ�����ֲ������ʷ�е������Σ�������ռ��������ʷ�����ƣ�������ֻ�Ǹ����ڶ����������棬���DZ���ֲ�������ֲ������ʷ�Ĺ�ͬ�ص㡣������ֲ��������������ֲ��ĸ����˻������������Ϊ���ӡ��������������Σ��������������͵������������Σ���������������������ʷ���й���ؽ�����ֵ�����Ϊ�������棨alternationofgeneration����
��������ֲ�����������г��ֵļ������Ѻ��ܾ����ã������ںϣ�������������ʷ�Ĺؼ���Ҳ���������������ת�۵㣬�����������ӡ�����ֲ�����������ģʽͼ�ͼ���������ͼ�⣬��ͼ4-80��
��
 ��
����ϰ˼����
��
����1��ֲ��ķ�ֳ������Щ��Ҫ������ѧ���壿ֲ��ķ�ֳ�ɷ�Ϊ�ļ������ͣ����ַ�ֳ���͵��ص���ʲô��
����2��ʲô����ȻӪ����ֳ���ٸ�����ȻӪ����ֳ��ʵ������˵����
����3��ʲô���˹�Ӫ����ֳ�������������õ��˹�Ӫ����ֳ���ļ��֣��˹�Ӫ����ֳ�������ϵ�����������ʲô��
����4��Ǥ���ѹ����ֳʱ��ע����Щ��Ҫ�Ļ��ڣ����ߵľ�������кβ�ͬ��
����5���ӷ�ֳ��Ǥ�塢ѹ���кβ�ͬ�����Լӷ�ֳ����Ǥ���ѹ����ֳ��
����6���ӷ�ֳ�ijɹ��ؼ���ʲô������IJ����������ļ��֣�
����7���������������ģ�ֲ����Ӫ������ת����ֳ��������ֲ���Ϸ��������������仯��
����8�����͵Ļ�����Щ��Ҫ���֣������ֵ���̬�ͽṹ��Σ�
����9�������ڲ�ֲͬ��������ı仯�����Σ�ʲô��ͬ�����������������������ޱ���������˵����
����10�������ͻ�����롢�������Σ�
����11����ֲ��ϵͳ�ݻ������У����ĸ���ɲ����������������ݻ����ƶ��仯�ģ���Щ�仯�Ƿ���ͬ����չ�ģ�����˵����
����12�����е���̬�仯���ʹ�ӷ��ͻ���������ɲ��ֵ�λ��Ҳ��Ӧ������仯���ɴ˶�����IJ�ͬ�ӷ�λ�õĻ���������ʲô��
����13��˵��һ��С�Ľṹ��
����14����ʽ�ͻ���ʽ�ĺ�����ʲô����λ��ƻ�ͼʽ��дһ������ʽ������˵����
����15��ʲô�ǻ�������������Ҫ������ʲô������˵����������Ҫ������
����16����ҩ�ڵķ���������Σ�ҩ���е���ձ����С�����γɹ���������ʲô��Ҫ�����ã�
����17������ԭϸ������ΪС���ӵĹ�����Σ����컨�۵�һ��ṹ��Σ�
����18��ʲô�ǻ��۰��������Բ��������������������ԭ����ʲô��
����19������ķ���������Σ����ļ������ͣ�
����20��ʲô��̥�������ʶ�����̥�����ͣ�
����21�������͡�˫���ͺ��İ��͵����ҷ���������γɵģ�
����22������8�ˡ�7ϸ�����ҵķ��������������ģ� 7ϸ�����Ҹ�ϸ�������ƺ�������Σ�
����23��ʲô���Ի����ۺ��컨���ۣ��컨���۱��Ի������ں���ķ��������и�����Խ�ԣ�ԭ����ʲô��ֲ������ڻ�������̬�ṹ����ʽ��������Ի����۵ķ������Ի�����������Ȼ�类����������ԭ������ʲô��
����24�����ֲ�ͬ���۷�ʽ������̬�ṹ������Σ�
����25������ֲ���˫�ܾ����̼������塣
����26����������ͷ�ϵ��ȷ���Ϊʲô������ͺͲ��͵�����ϸ����֯ѧ�ĽǶȿ����ͻ��͵�ԭ����ʲô��
����27��ʲô�����ں���ֳ�Ͷ�������
����28�����˺�С���ߵķ���������Σ������к���ͬ�㣿
����29���������鷢�����͵���ϸ������Σ�
����30����Ƥ�Ľṹ������Σ���Ƥ�����ʡ������ʵ�Ĺ�Ƥ֮���Ƿ���һ��������ԣ�
����31��ʲô�ǵ��Խ�ʵ���Է����յ����Խ�ʵ���������к���Ҫ���壿
32���оٸ����ʵ�Ľṹ������
33�����Ӻ�ʵ�Ĵ�������Щ��ʽ��
34����˳����ֲ�������ʷ���ñ��г�����ʷ�и����εķ�չ˳������С�����ҵķ�������С���ӵ��γɣ��ơ���������ķ������ơ������ӵ��γɣ��ܾ����̣����ܾ�������Ĺ�ʵ����һ���ij��塣ע�����εĺ���仯��
